Sangaran Pushpa Gandi, Ibrahim Zaridatul Aini, Chik Zamri, Mohamed Zahurin, Ahmadiani Abolhassan
Department of Pharmacology, Faculty of Medicine, University of Malaya, Kuala Lumpur, Malaysia.
Department of Pharmacology, School of Medicine, Shahid Beheshti University of Medical Sciences, Evin, Tehran, Iran.
Front Cell Neurosci. 2021 Jan 22;14:598453. doi: 10.3389/fncel.2020.598453. eCollection 2020.
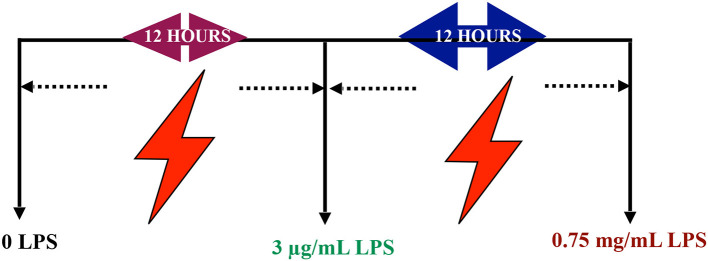
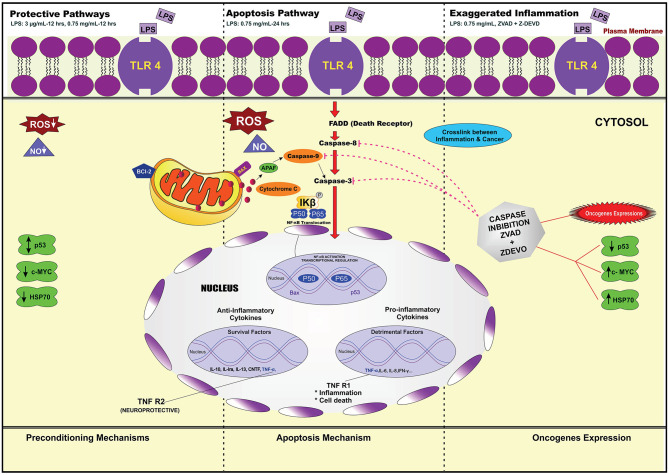
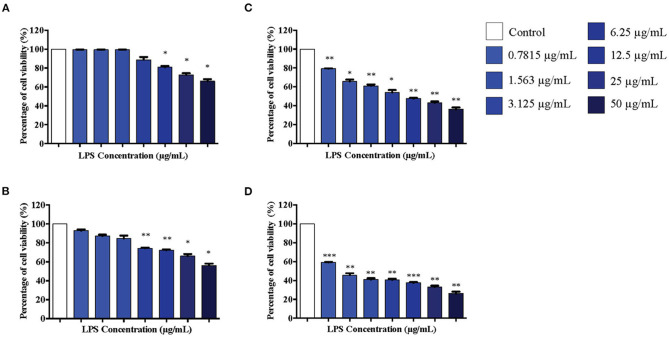
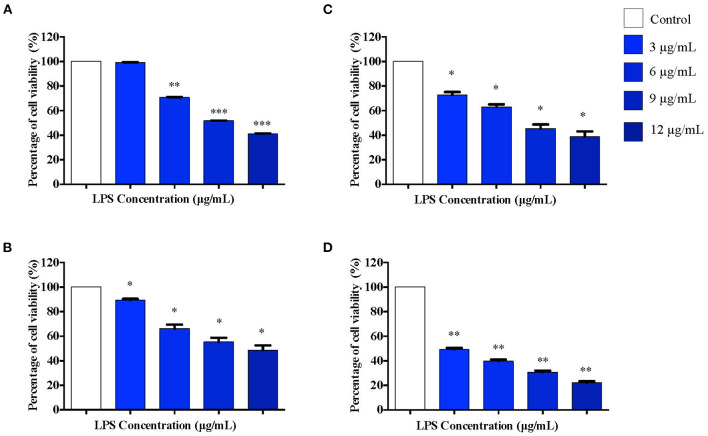
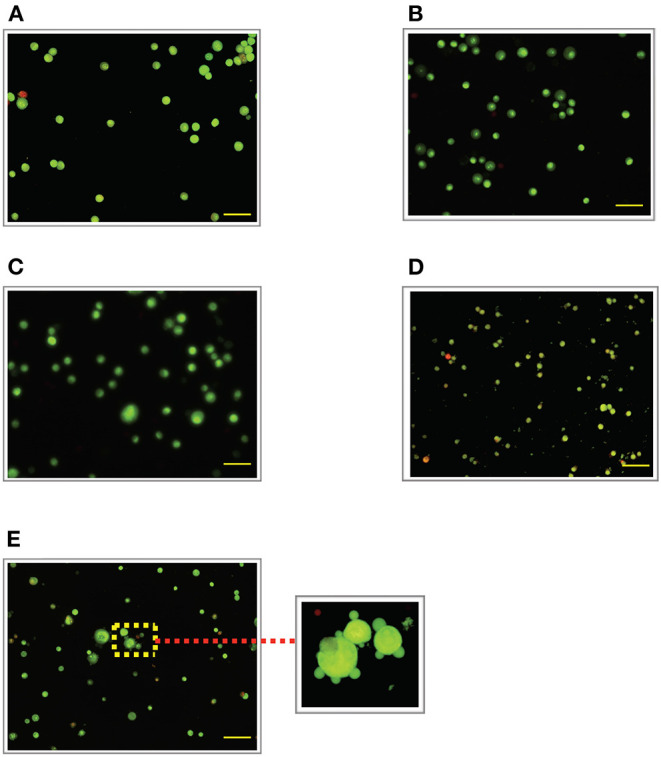
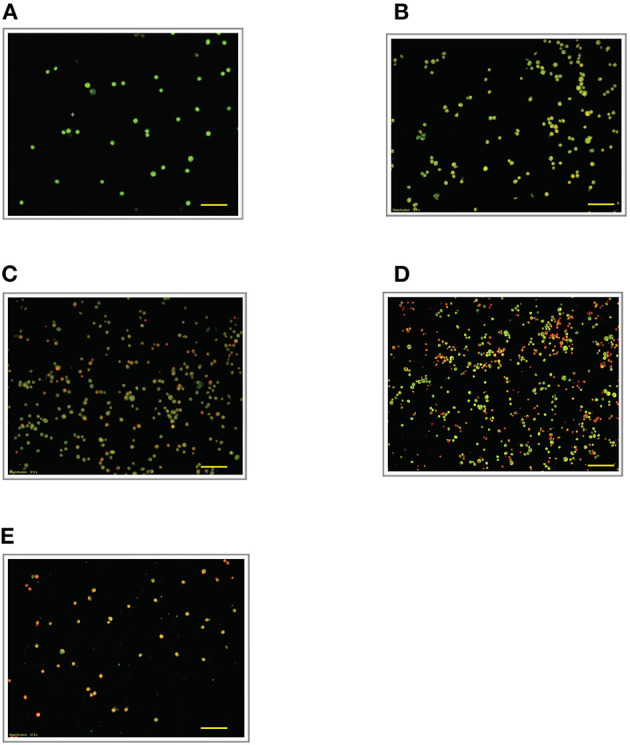
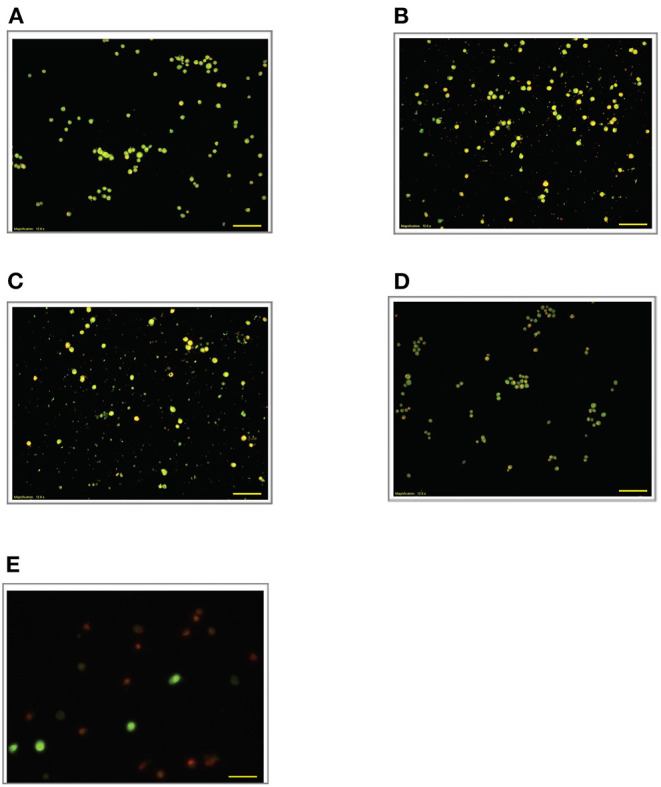
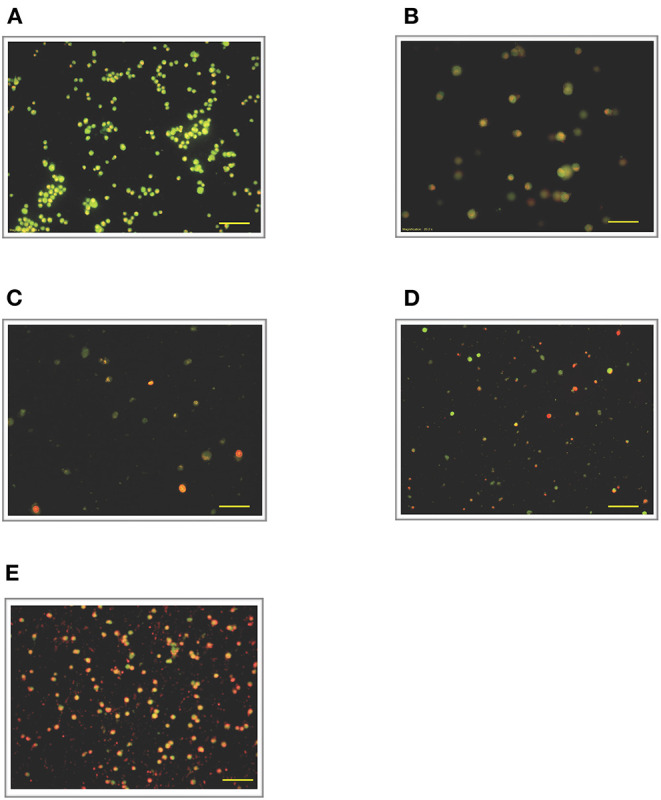
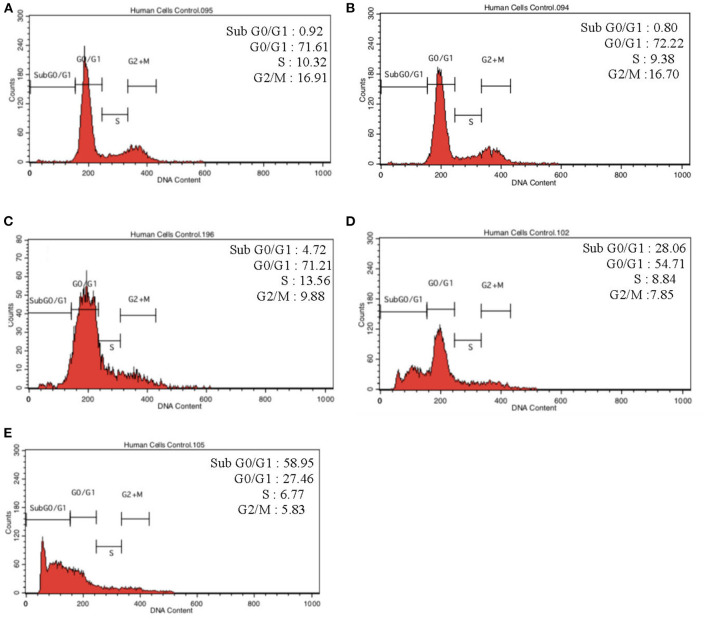
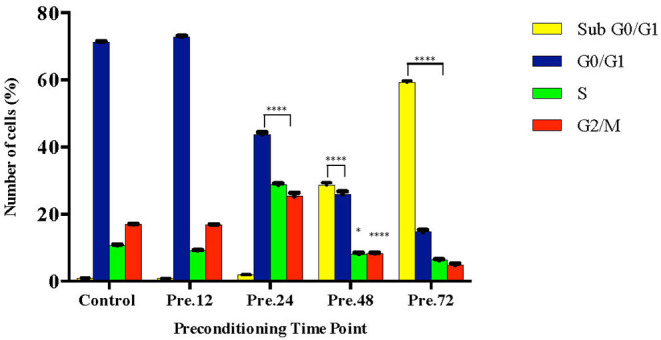
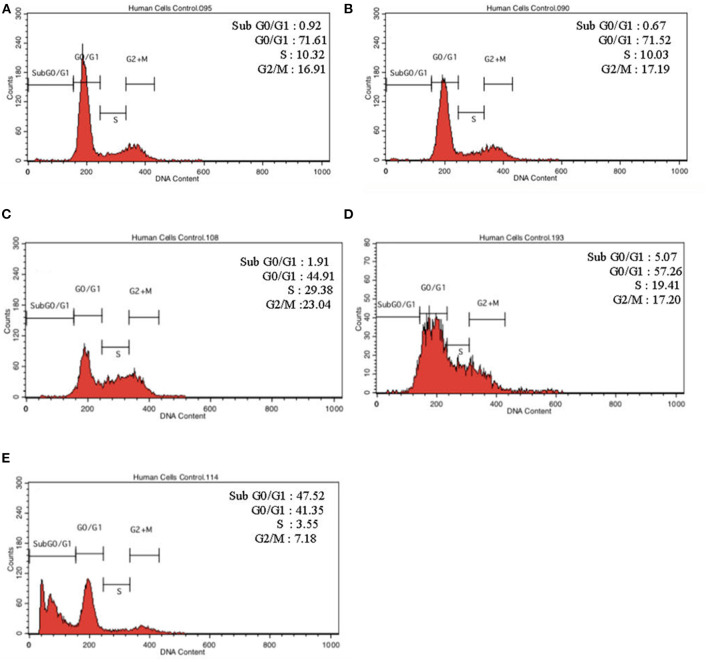
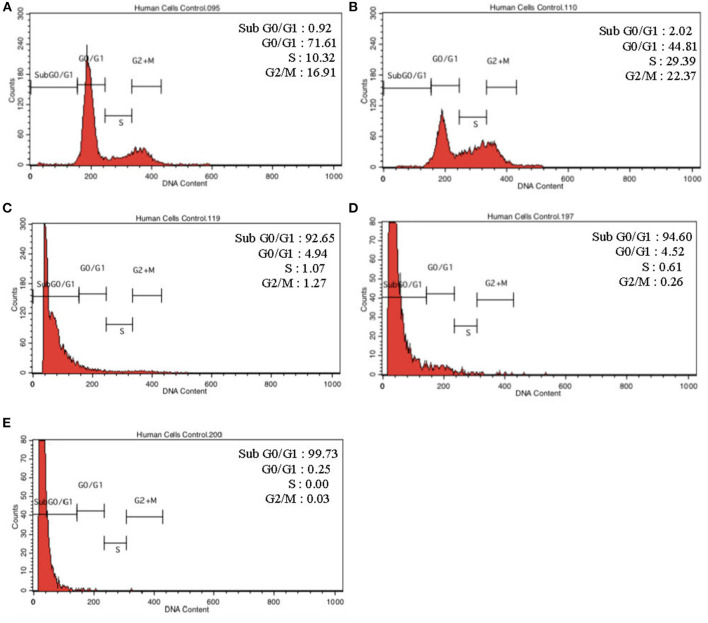
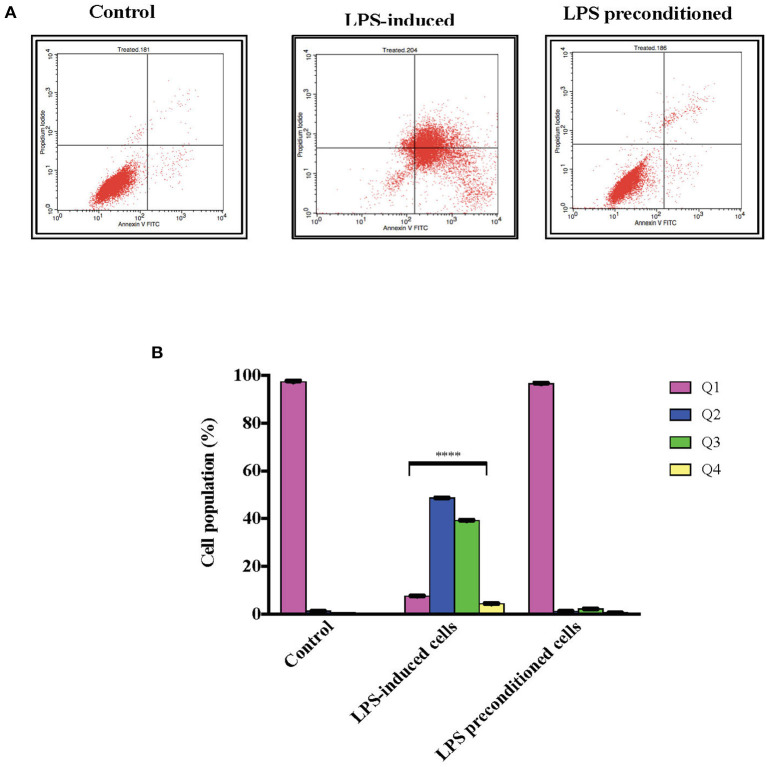
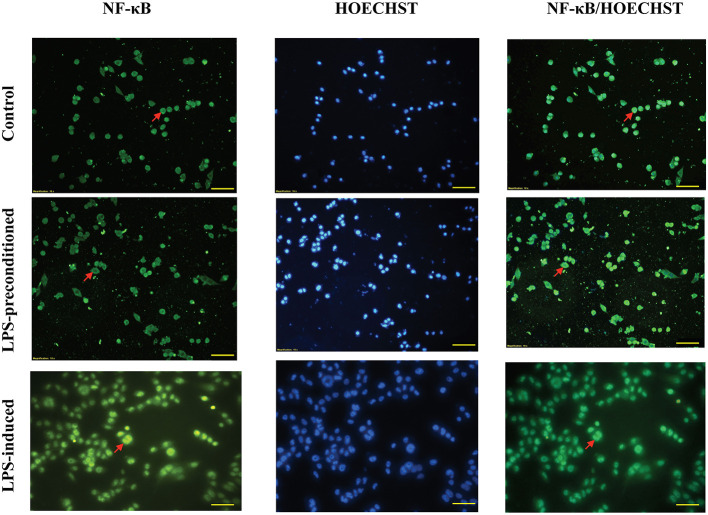
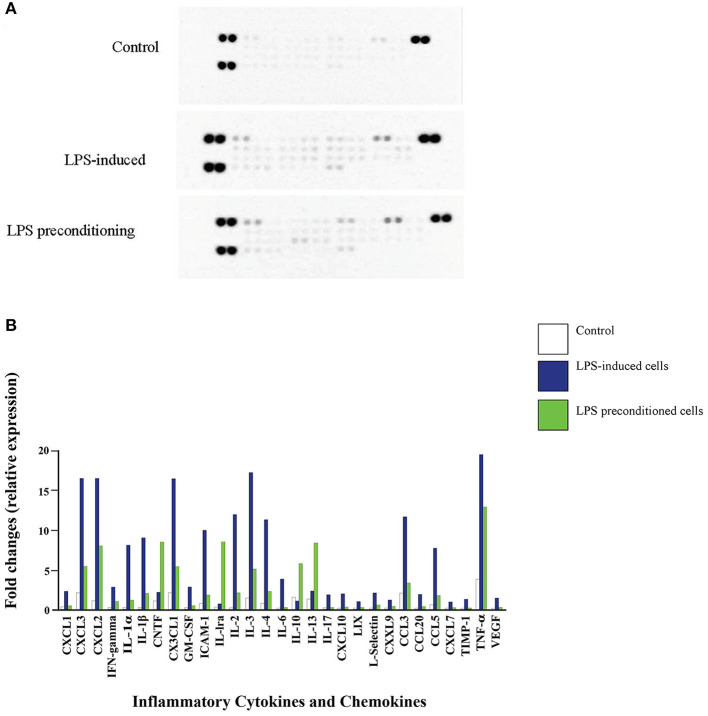
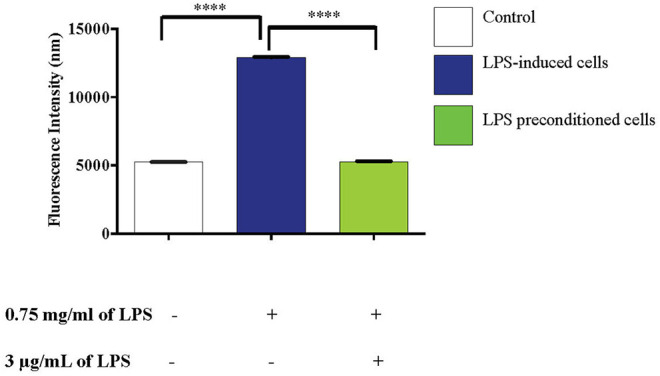
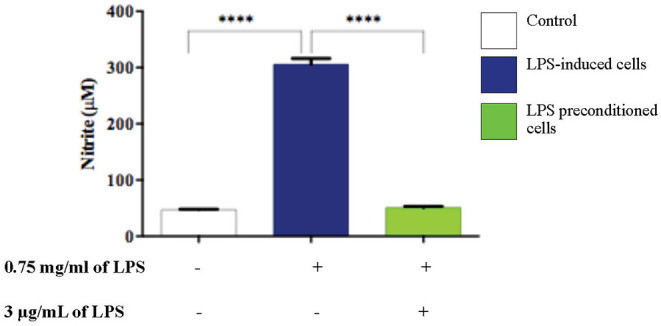
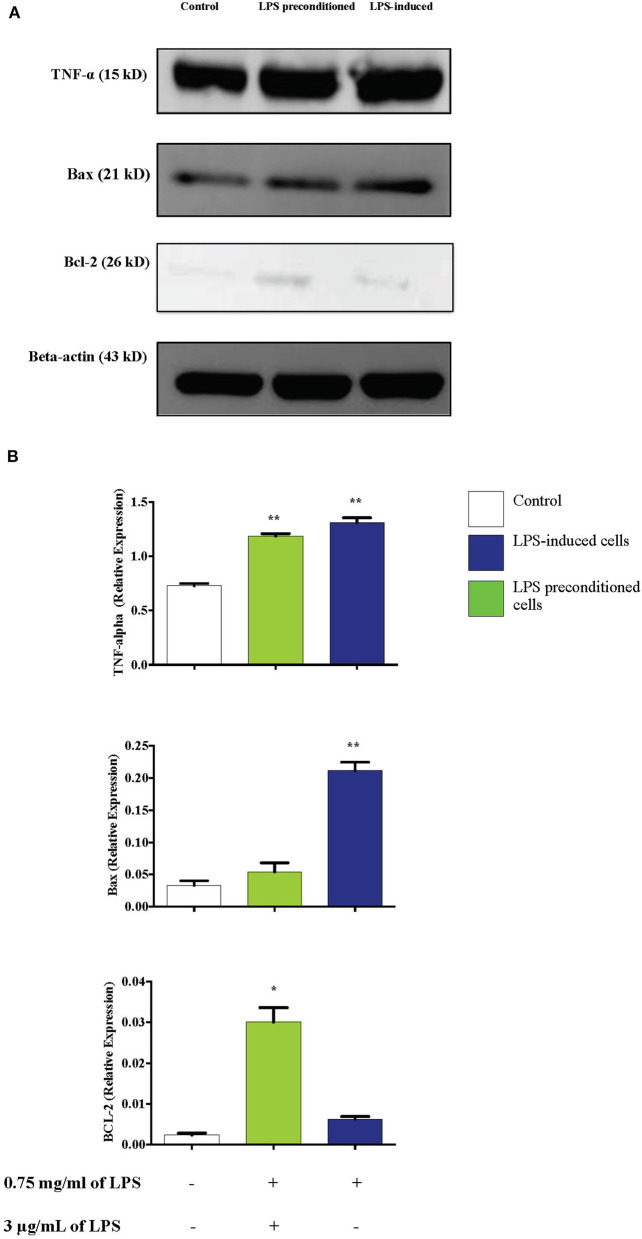
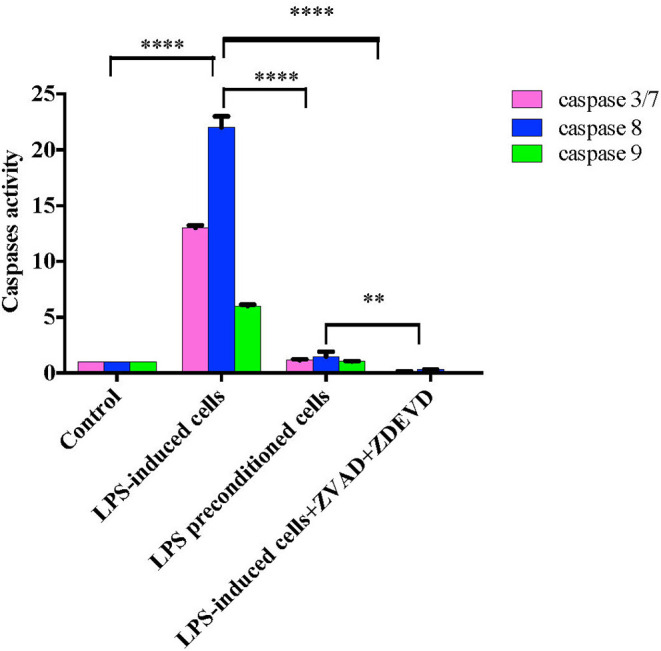
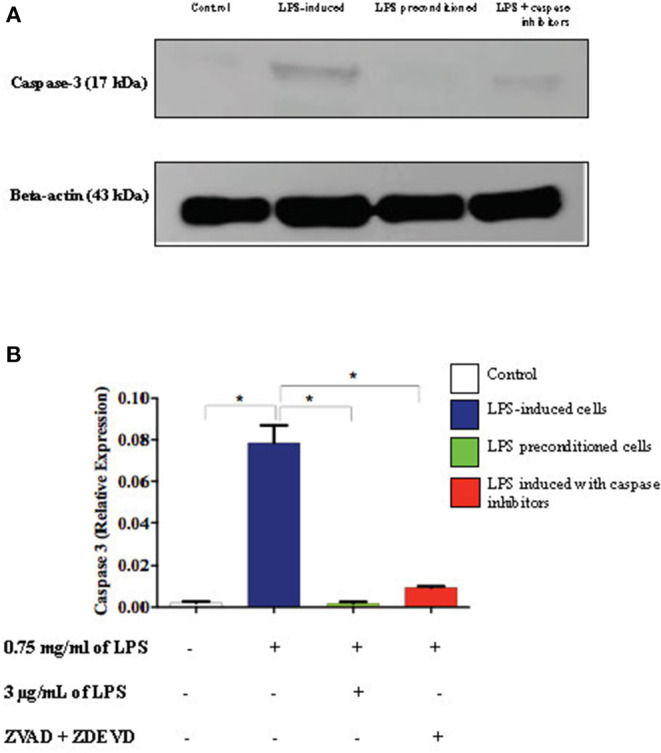
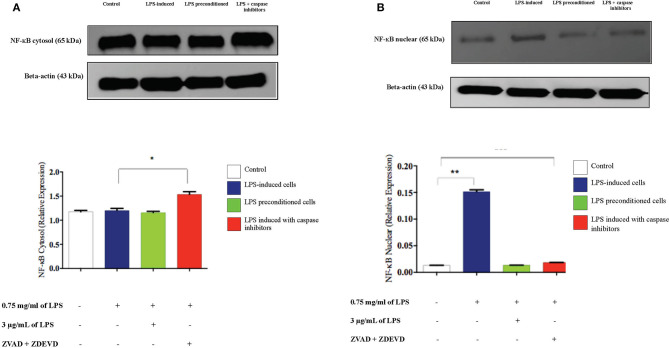
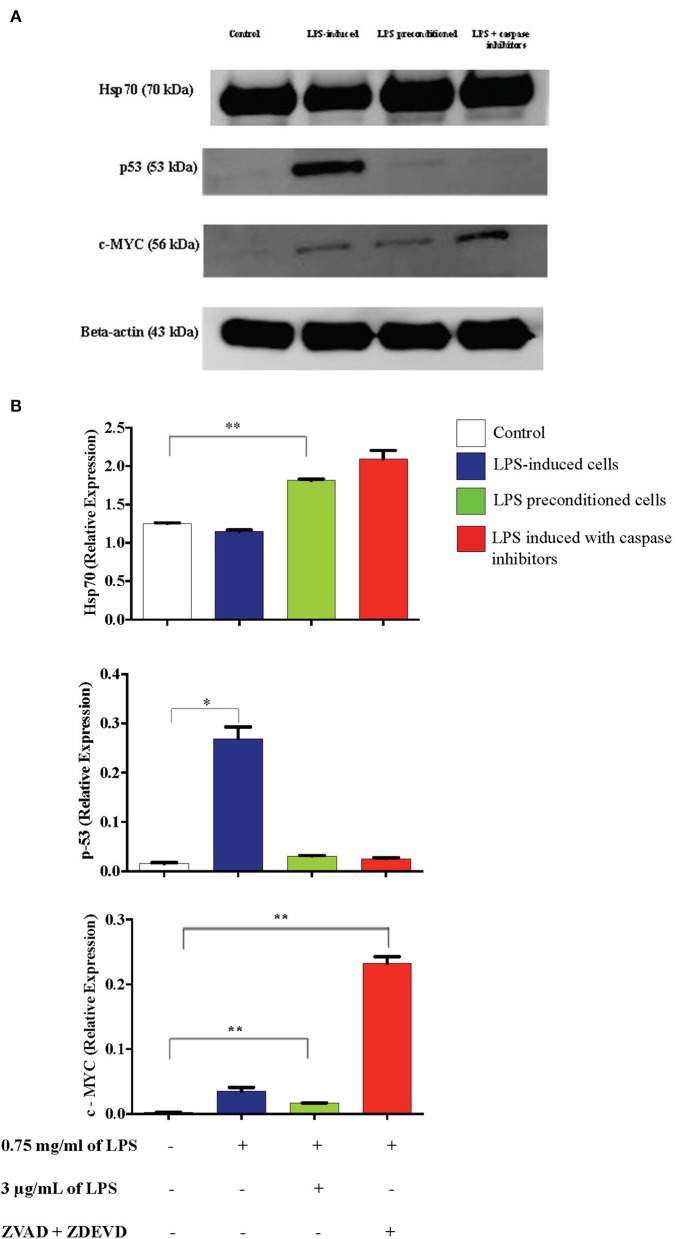
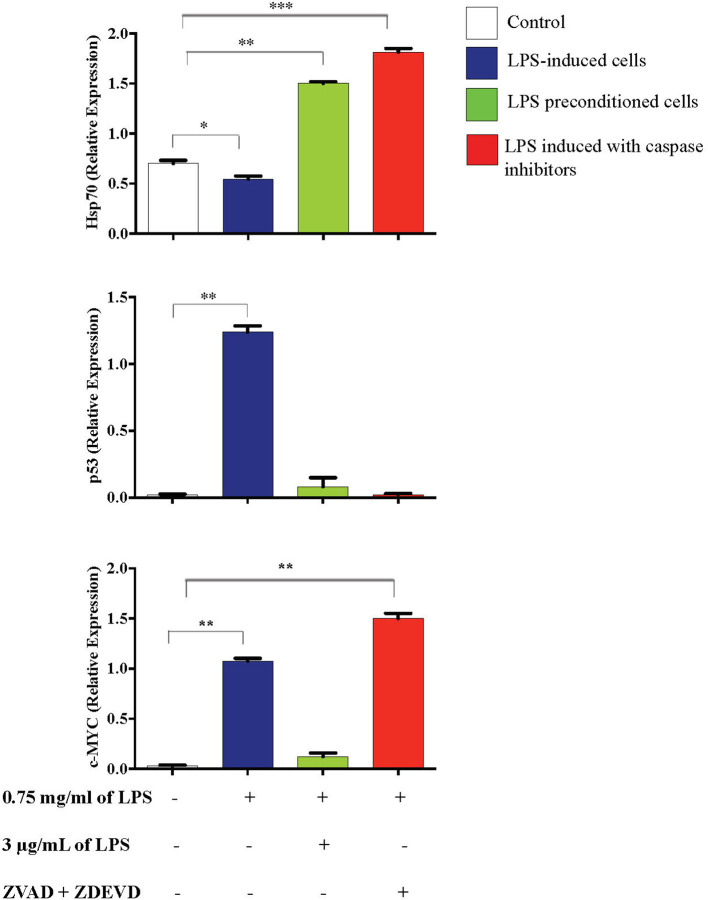
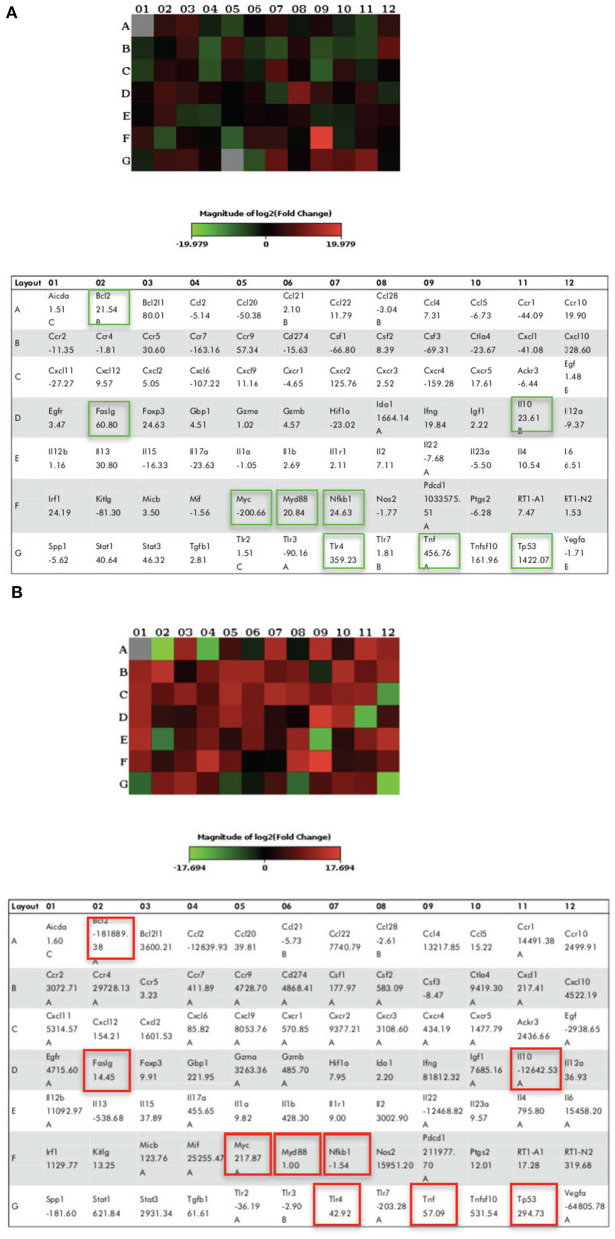
Lipopolysacharide (LPS) pre-conditioning (PC), has been shown to exert protective effects against cytotoxic effects. Therefore, we hypothesized, the tolerance produced by LPS PC will be resulted by the alterations and modifications in gene and protein expression. With reference to the results of MTT assays, AO/PI staining, and Annexin V-FITC analyses of LPS concentration (0.7815-50 μg/mL) and time-dependent (12-72 h) experiments, the pre-exposure to 3 μg/mL LPS for 12 h protected the differentiated PC12 cells against 0.75 mg/mL LPS apoptotic concentration. LPS-treated cells secreted more inflammatory cytokines like IL-1α, IL-1β, IL-2, IL-3, IL-4, IL-6, IL-17, IFN-γ, and TNF-α than LPS-PC cells. The production of inflammatory mediators ROS and NO was also higher in the LPS-induced cells compared to LPS-PC cells. Conversely, anti-inflammatory cytokines (like IL-10, IL-13, CNTF, and IL-1Ra) were upregulated in the LPS-PC cells but not in the LPS-induced cells. Meanwhile, the LPS initiated caspase-8 which in turn activates effector caspase 3/7. When the activities of caspases in the LPS-induced cells were inhibited using z-VADfmk and z-DEVDfmk, the expressions of c-MYC and Hsp70 were increased, but p53 was reduced. The potential molecules associated with protective and destructive effect was measured by RT Profiler PCR array to elucidate the signaling pathways and suggested inhibition NF-κB/caspase-3 signaling pathway regulates the cytoprotective genes and proto-oncogenes. In conclusion, this study provides a basis for future research to better understand the molecular mechanism underlying LPS pre-conditioning /TLR4 pre-activation and its functional role in offering cytoprotective response in neuronal environment.
脂多糖(LPS)预处理(PC)已被证明对细胞毒性作用具有保护作用。因此,我们推测,LPS PC产生的耐受性将由基因和蛋白质表达的改变和修饰所致。参照MTT分析、AO/PI染色以及LPS浓度(0.7815 - 50μg/mL)和时间依赖性(12 - 72小时)实验的Annexin V-FITC分析结果,预先暴露于3μg/mL LPS 12小时可保护分化的PC12细胞免受0.75mg/mL LPS凋亡浓度的影响。与LPS-PC细胞相比,LPS处理的细胞分泌更多的炎性细胞因子,如IL-1α、IL-1β、IL-2、IL-3、IL-4、IL-6、IL-17、IFN-γ和TNF-α。与LPS-PC细胞相比,LPS诱导的细胞中炎性介质ROS和NO的产生也更高。相反,抗炎细胞因子(如IL-10、IL-13、CNTF和IL-1Ra)在LPS-PC细胞中上调,但在LPS诱导的细胞中未上调。同时,LPS启动了半胱天冬酶-8,进而激活效应半胱天冬酶3/7。当使用z-VADfmk和z-DEVDfmk抑制LPS诱导细胞中的半胱天冬酶活性时,c-MYC和Hsp70的表达增加,但p53减少。通过RT Profiler PCR阵列检测与保护和破坏作用相关的潜在分子,以阐明信号通路,并提示抑制NF-κB/半胱天冬酶-3信号通路调节细胞保护基因和原癌基因。总之,本研究为未来更好地理解LPS预处理/TLR4预激活的分子机制及其在神经元环境中提供细胞保护反应的功能作用提供了基础。