Department of Biochemistry and Molecular Genetics, University of Colorado Anschutz Medical Campus, Aurora, CO, USA.
Telomere Biology Laboratory, Laboratory of Biochemistry and Molecular Biology, Center for Cancer Research, NCI, NIH, Bethesda, MD, USA.
Nature. 2021 Mar;591(7851):671-676. doi: 10.1038/s41586-021-03279-8. Epub 2021 Mar 3.
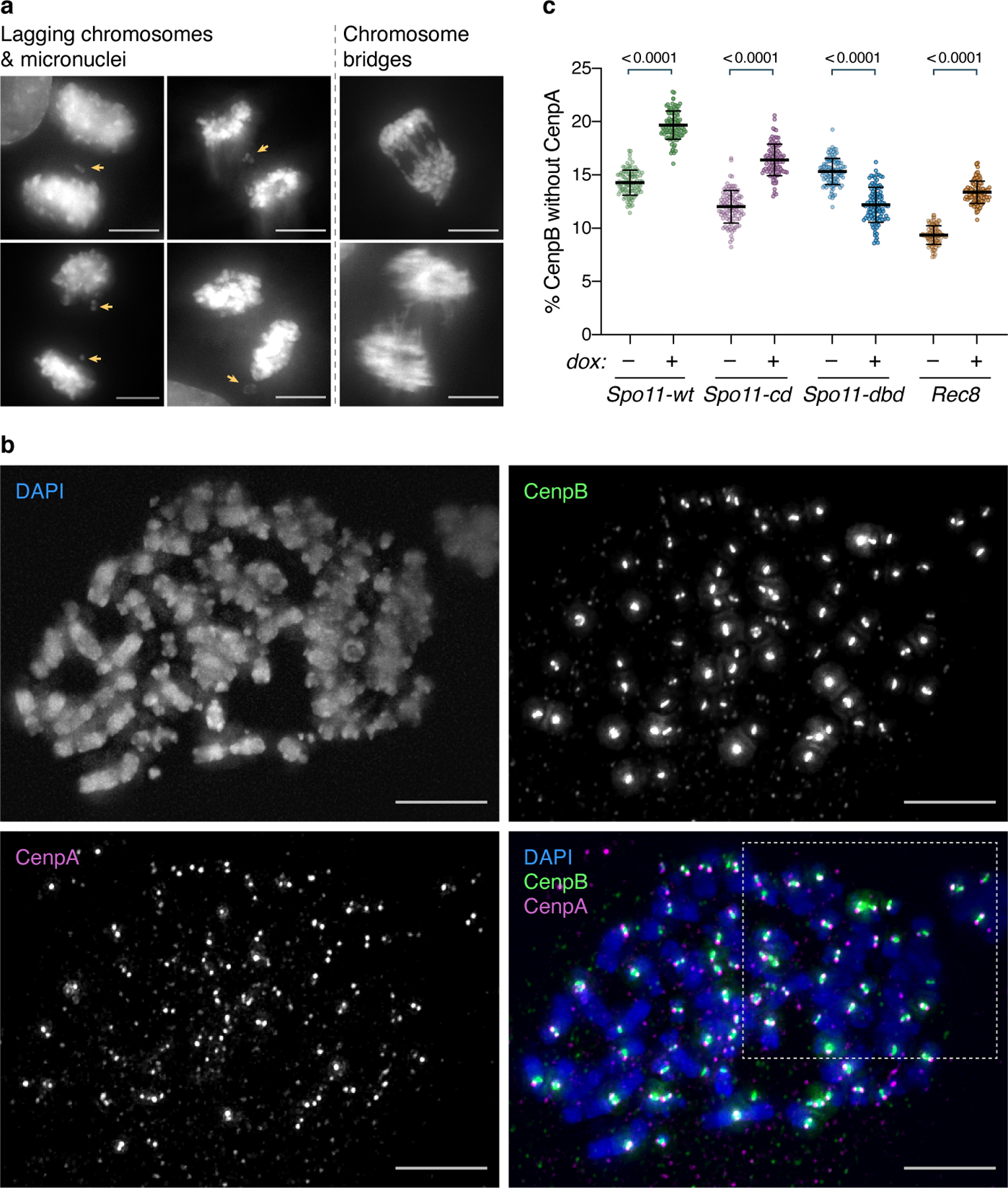
Meiotic processes are potentially dangerous to genome stability and could be disastrous if activated in proliferative cells. Here we show that two key meiosis-defining proteins, the topoisomerase Spo11 (which forms double-strand breaks) and the meiotic cohesin Rec8, can dismantle centromeres. This dismantlement is normally observable only in mutant cells that lack the telomere bouquet, which provides a nuclear microdomain conducive to centromere reassembly; however, overexpression of Spo11 or Rec8 leads to levels of centromere dismantlement that cannot be countered by the bouquet. Specific nucleosome remodelling factors mediate centromere dismantlement by Spo11 and Rec8. Ectopic expression of either protein in proliferating cells leads to the loss of mitotic kinetochores in both fission yeast and human cells. Hence, while centromeric chromatin has been characterized as extraordinarily stable, Spo11 and Rec8 challenge this stability and may jeopardize kinetochores in cancers that express meiotic proteins.
减数分裂过程对基因组稳定性有潜在的危险,如果在增殖细胞中激活,可能会造成灾难性的后果。在这里,我们表明,两种关键的减数分裂定义蛋白,拓扑异构酶 Spo11(形成双链断裂)和减数分裂黏合蛋白 Rec8,可以拆解着丝粒。这种拆解通常只能在缺乏端粒花束的突变细胞中观察到,端粒花束提供了一个有利于着丝粒重新组装的核微域;然而,Spo11 或 Rec8 的过表达会导致无法通过花束来抵消的着丝粒拆解水平。特定的核小体重塑因子通过 Spo11 和 Rec8 介导着丝粒的拆解。在增殖细胞中异位表达这两种蛋白中的任何一种,都会导致裂殖酵母和人类细胞中有丝分裂动粒的丢失。因此,虽然着丝粒染色质已经被描述为具有极高的稳定性,但 Spo11 和 Rec8 挑战了这种稳定性,并且可能会危及表达减数分裂蛋白的癌症中的动粒。