Department of Parasitology, Faculty of Science, Charles University, BIOCEV, Průmyslová 595, 25250, Vestec, Czech Republic.
Department of Cellular Biology, University of Georgia, Athens, GA, USA.
BMC Ecol Evol. 2021 Mar 19;21(1):46. doi: 10.1186/s12862-021-01777-x.
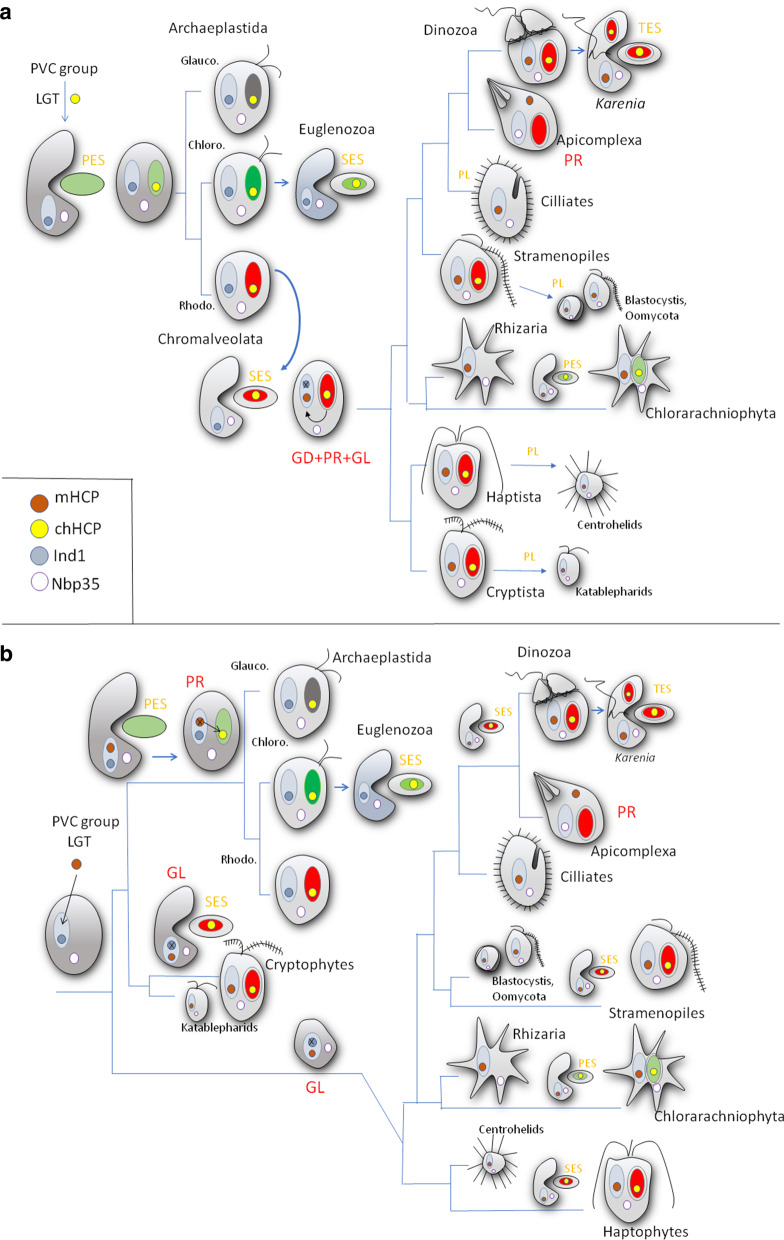
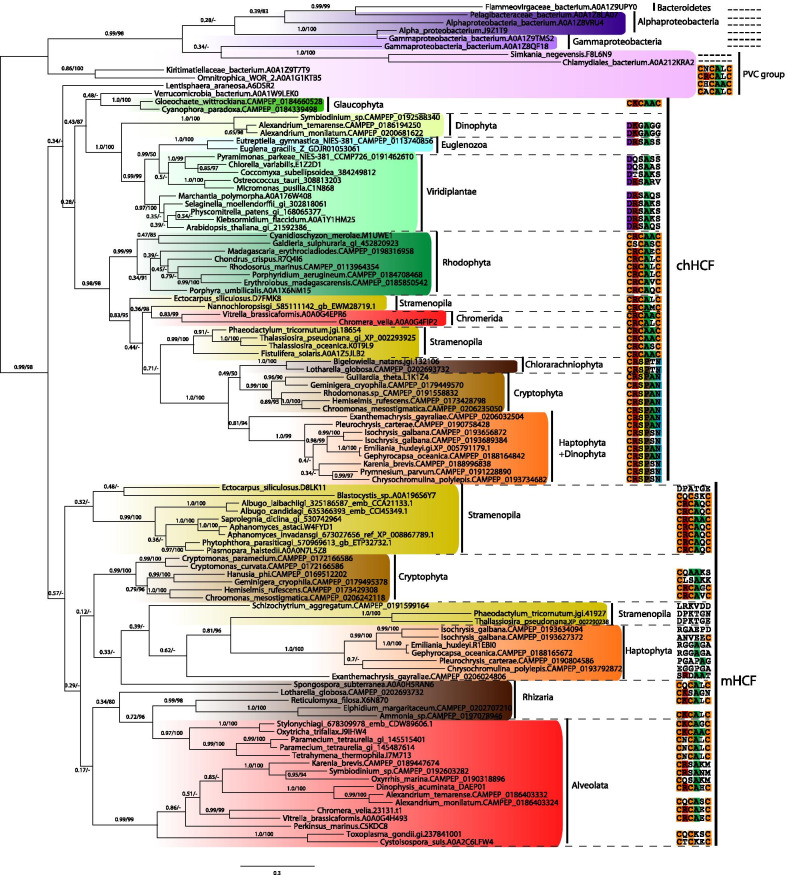
Nbp35-like proteins (Nbp35, Cfd1, HCF101, Ind1, and AbpC) are P-loop NTPases that serve as components of iron-sulfur cluster (FeS) assembly machineries. In eukaryotes, Ind1 is present in mitochondria, and its function is associated with the assembly of FeS clusters in subunits of respiratory Complex I, Nbp35 and Cfd1 are the components of the cytosolic FeS assembly (CIA) pathway, and HCF101 is involved in FeS assembly of photosystem I in plastids of plants (chHCF101). The AbpC protein operates in Bacteria and Archaea. To date, the cellular distribution of these proteins is considered to be highly conserved with only a few exceptions.
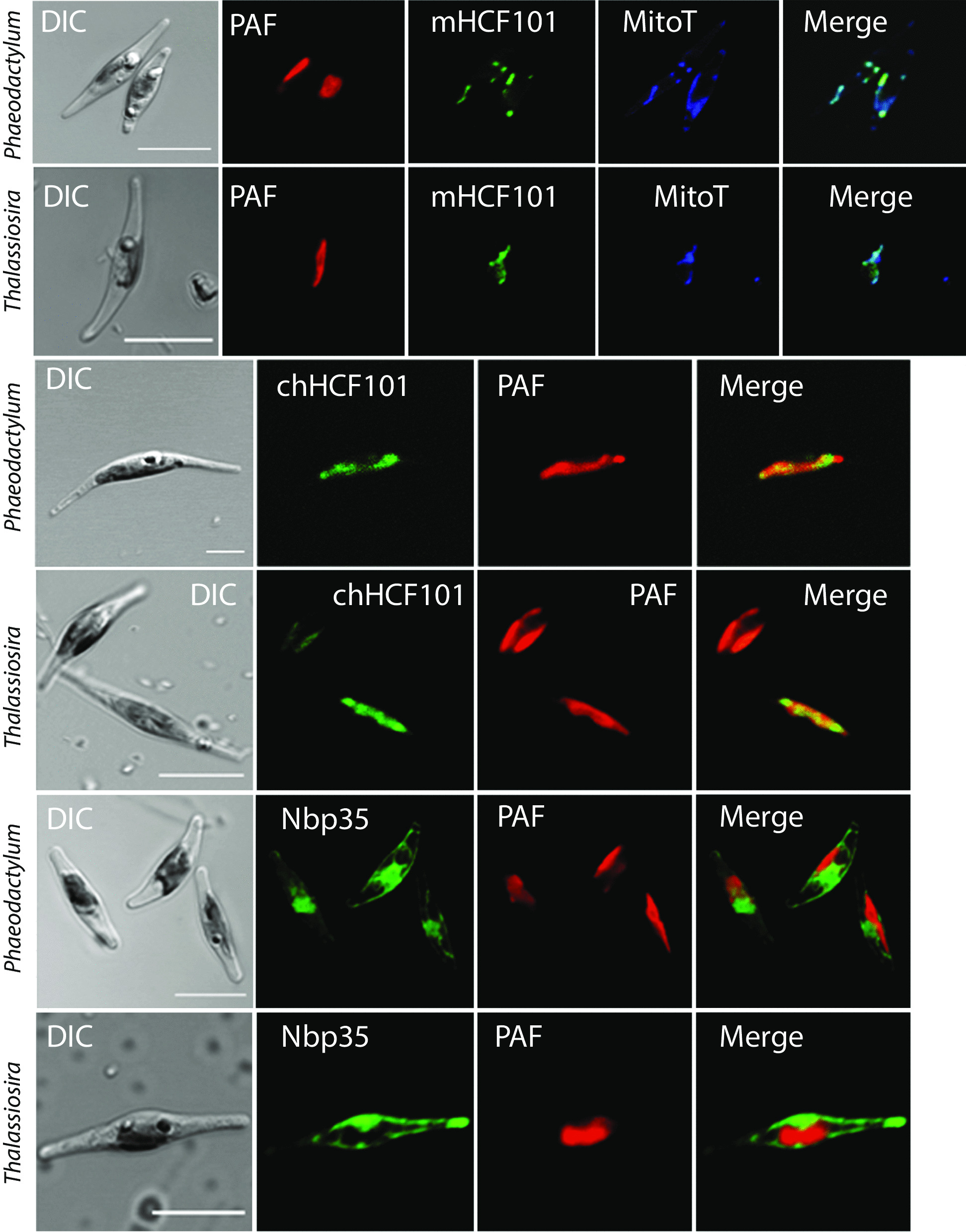
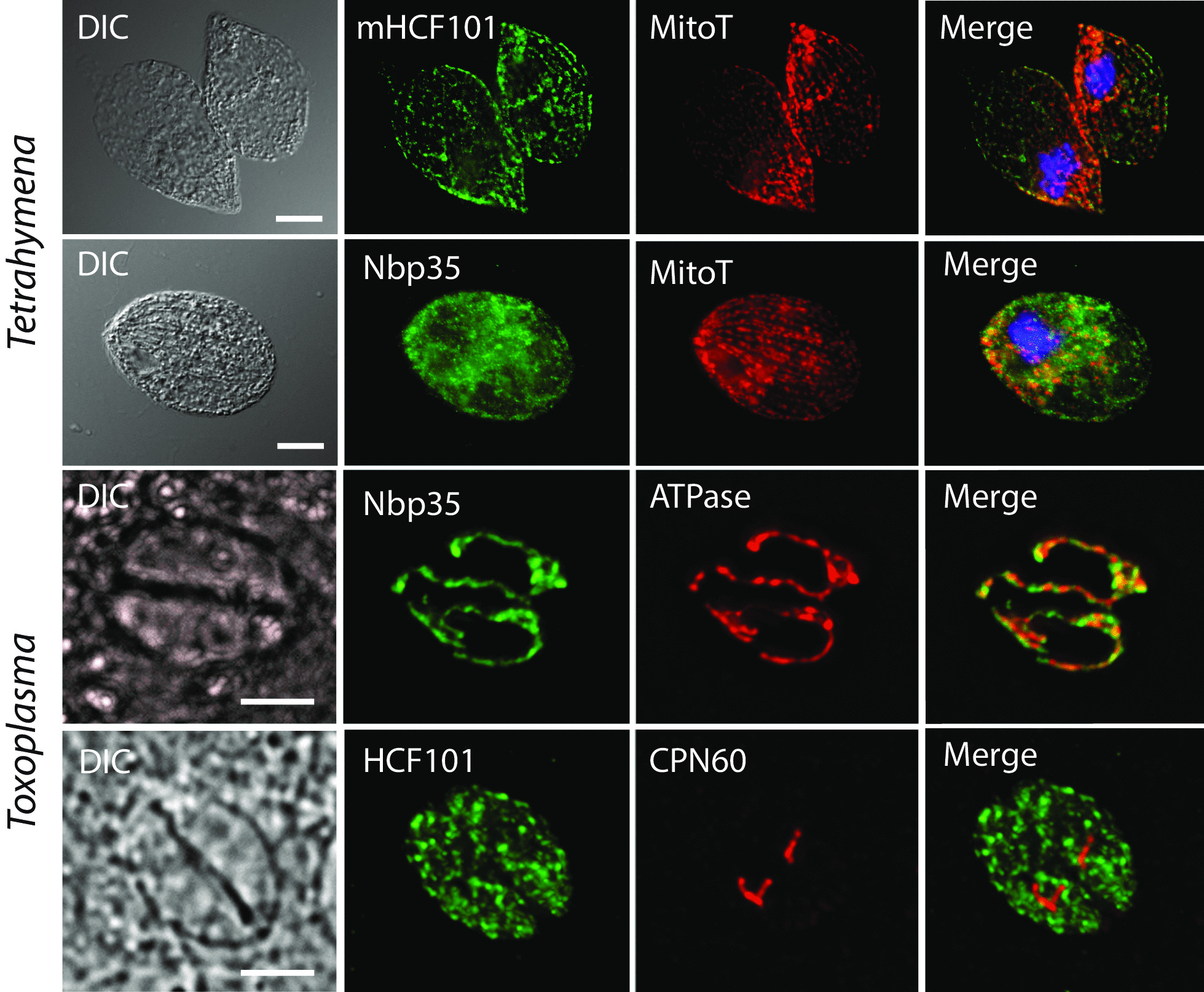
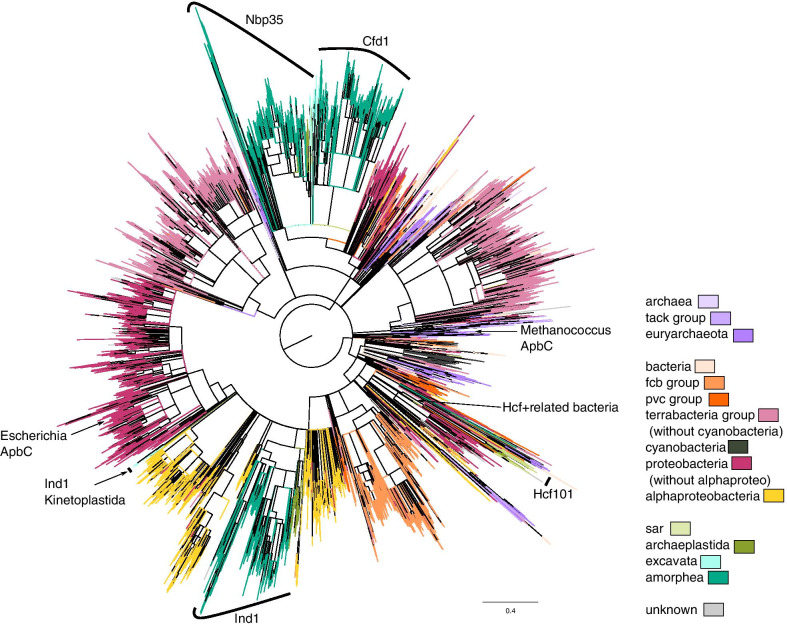
We searched for the genes of all members of the Nbp35-like protein family and analyzed their targeting sequences. Nbp35 and Cfd1 were predicted to reside in the cytoplasm with some exceptions of Nbp35 localization to the mitochondria; Ind1was found in the mitochondria, and HCF101 was predicted to reside in plastids (chHCF101) of all photosynthetically active eukaryotes. Surprisingly, we found a second HCF101 paralog in all members of Cryptista, Haptista, and SAR that was predicted to predominantly target mitochondria (mHCF101), whereas Ind1 appeared to be absent in these organisms. We also identified a few exceptions, as apicomplexans possess mHCF101 predicted to localize in the cytosol and Nbp35 in the mitochondria. Our predictions were experimentally confirmed in selected representatives of Apicomplexa (Toxoplasma gondii), Stramenopila (Phaeodactylum tricornutum, Thalassiosira pseudonana), and Ciliophora (Tetrahymena thermophila) by tagging proteins with a transgenic reporter. Phylogenetic analysis suggested that chHCF101 and mHCF101 evolved from a common ancestral HCF101 independently of the Nbp35/Cfd1 and Ind1 proteins. Interestingly, phylogenetic analysis supports rather a lateral gene transfer of ancestral HCF101 from bacteria than its acquisition being associated with either α-proteobacterial or cyanobacterial endosymbionts.
Our searches for Nbp35-like proteins across eukaryotic lineages revealed that SAR, Haptista, and Cryptista possess mitochondrial HCF101. Because plastid localization of HCF101 was only known thus far, the discovery of its mitochondrial paralog explains confusion regarding the presence of HCF101 in organisms that possibly lost secondary plastids (e.g., ciliates, Cryptosporidium) or possess reduced nonphotosynthetic plastids (apicomplexans).
Nbp35 样蛋白(Nbp35、Cfd1、HCF101、Ind1 和 AbpC)是 P 环 NTP 酶,它们作为铁硫簇(FeS)组装机制的组成部分。在真核生物中,Ind1 存在于线粒体中,其功能与呼吸复合物 I 亚基中 FeS 簇的组装有关,Nbp35 和 Cfd1 是细胞质 FeS 组装(CIA)途径的组成部分,而 HCF101 参与植物质体(chHCF101)中光系统 I 的 FeS 组装。AbpC 蛋白存在于细菌和古菌中。迄今为止,这些蛋白质的细胞分布被认为是高度保守的,只有少数例外。
我们搜索了 Nbp35 样蛋白家族的所有成员的基因,并分析了它们的靶向序列。预测 Nbp35 和 Cfd1 位于细胞质中,但 Nbp35 定位于线粒体的情况除外;Ind1 存在于线粒体中,而 HCF101 预测存在于所有光合作用真核生物的质体(chHCF101)中。令人惊讶的是,我们在所有隐生生物界、原生动物界和 SAR 生物中发现了第二个 HCF101 旁系同源物,该蛋白主要定位于线粒体(mHCF101),而在这些生物中似乎不存在 Ind1。我们还发现了一些例外情况,例如顶复门生物拥有预测定位于细胞质的 mHCF101 和定位于线粒体的 Nbp35。我们通过用转基因报告蛋白标记选定的顶复门生物(刚地弓形虫)、Stramenopila(菱形藻、叉鞭金藻)和纤毛门生物(嗜热四膜虫)来验证我们的预测,实验结果证实了这一点。系统发育分析表明,chHCF101 和 mHCF101 是从共同的祖先 HCF101 进化而来的,与 Nbp35/Cfd1 和 Ind1 蛋白无关。有趣的是,系统发育分析支持祖先 HCF101 是从细菌水平转移而来的,而不是与 α-变形菌或蓝细菌内共生体的获得有关。
我们在真核生物谱系中搜索 Nbp35 样蛋白,发现 SAR、原生动物界和隐生生物界拥有线粒体 HCF101。由于此前只知道质体 HCF101 的定位,因此发现其线粒体旁系同源物解释了为何在可能失去次生质体(如纤毛虫、Cryptosporidium)或拥有非光合作用质体减少(顶复门生物)的生物中存在 HCF101 这一困惑。