School of Biological Sciences, Washington State University, Pullman, WA 99164-4236, USA.
Biocenter University of Cologne, Institute for Plant Science, Cologne 50674, Germany.
Plant Physiol. 2021 May 27;186(1):142-167. doi: 10.1093/plphys/kiaa117.
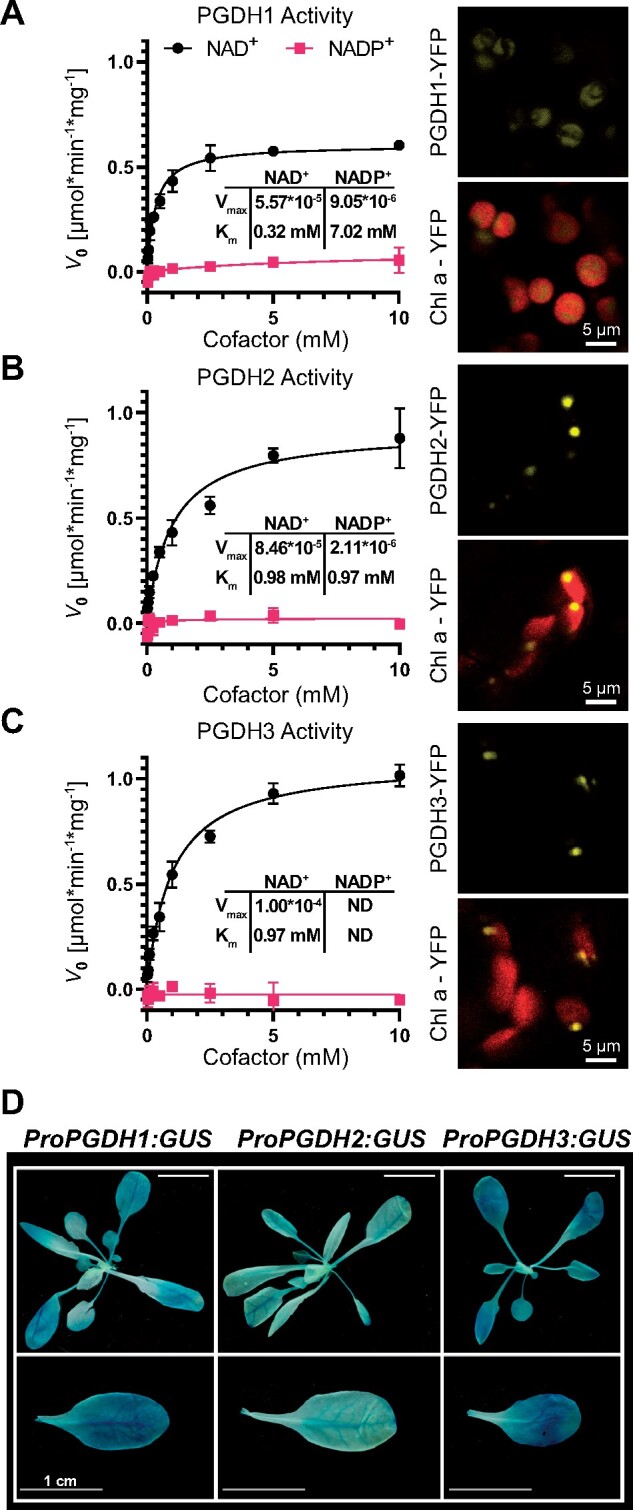
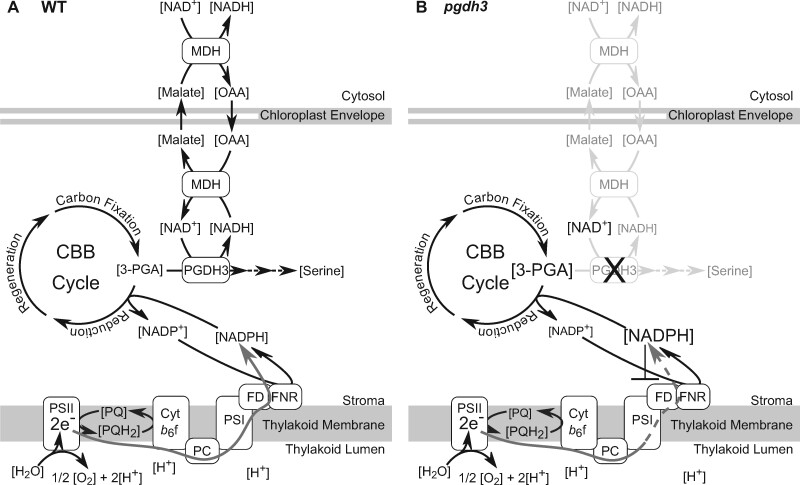
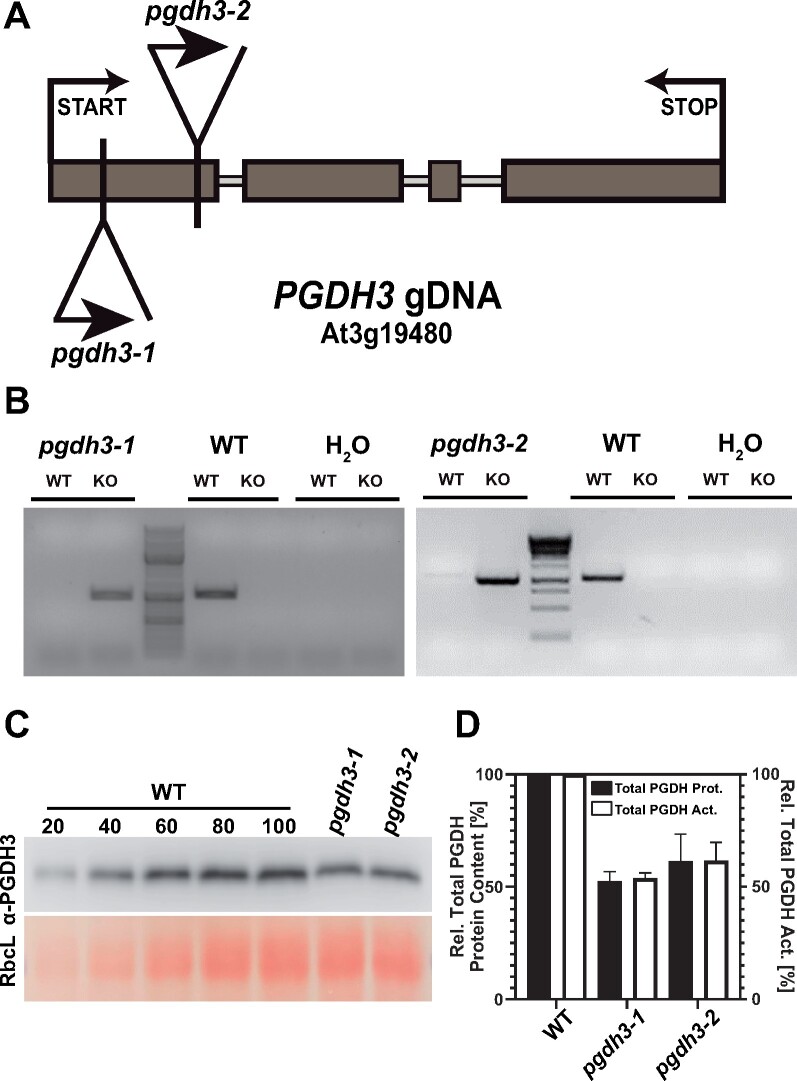
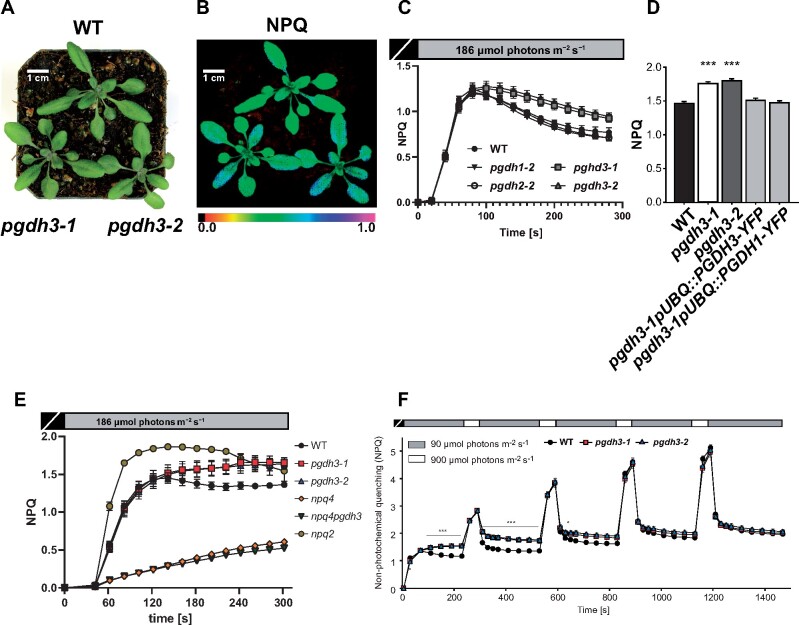
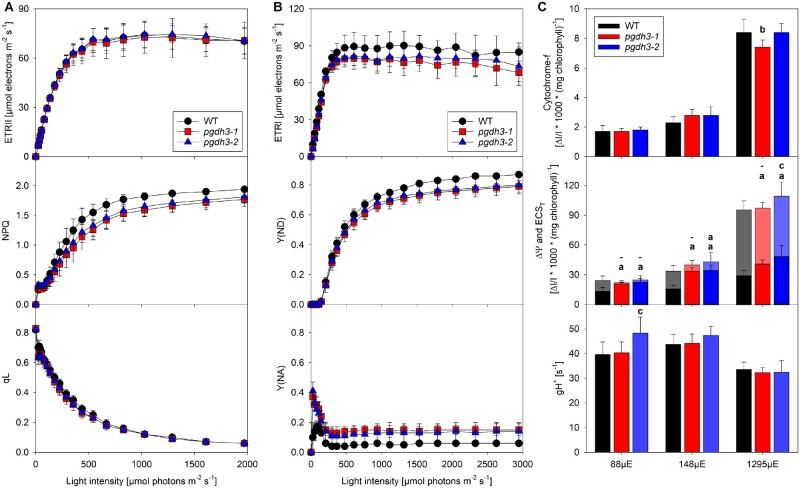
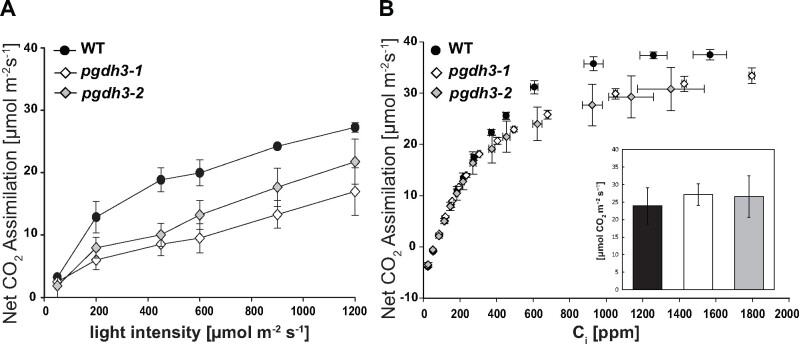
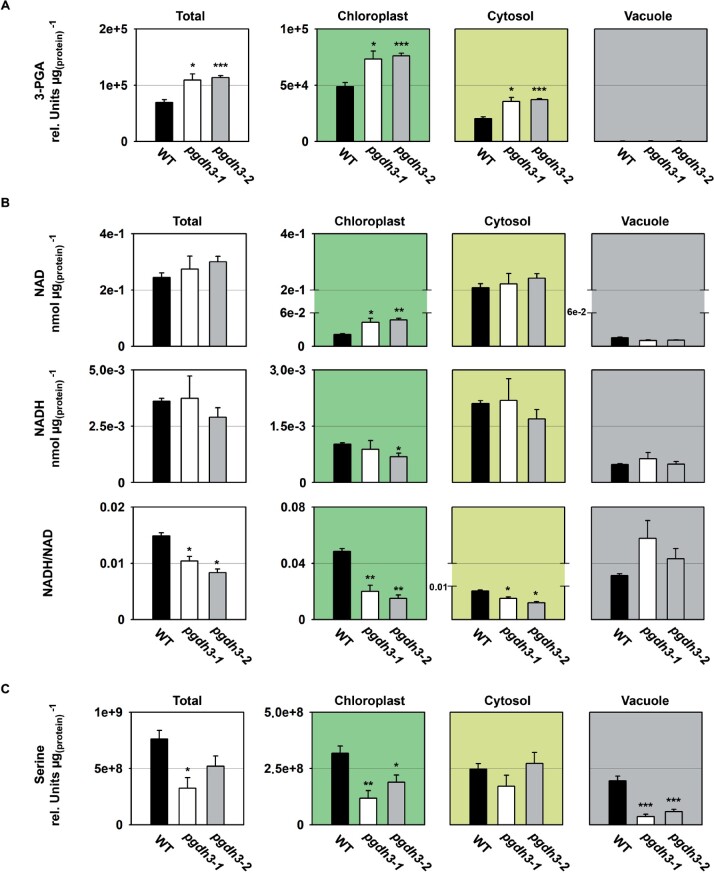
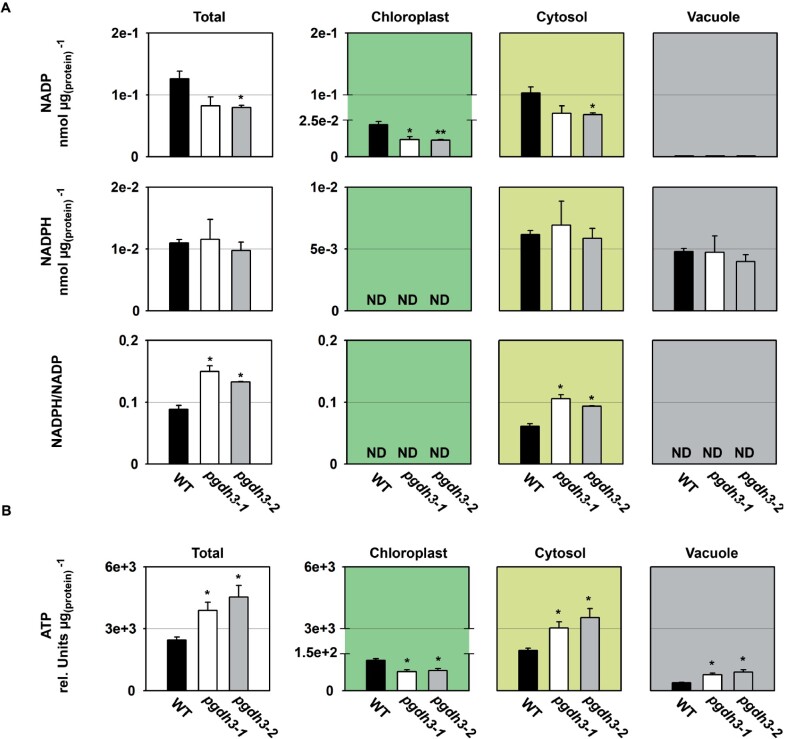
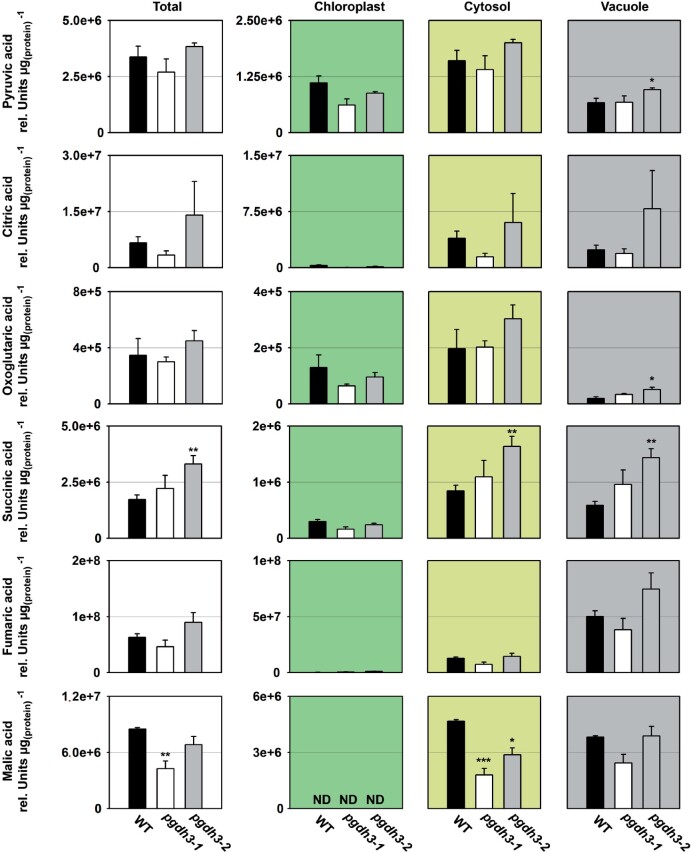
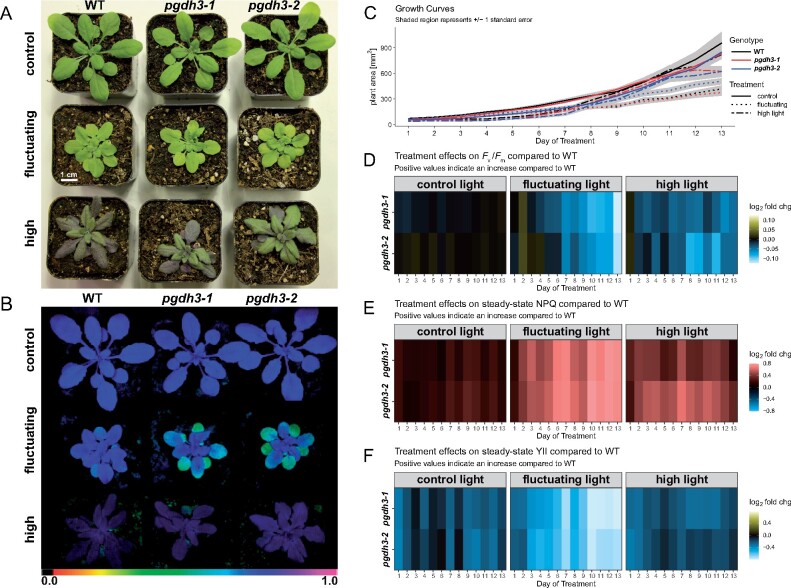
During photosynthesis, electrons travel from light-excited chlorophyll molecules along the electron transport chain to the final electron acceptor nicotinamide adenine dinucleotide phosphate (NADP) to form NADPH, which fuels the Calvin-Benson-Bassham cycle (CBBC). To allow photosynthetic reactions to occur flawlessly, a constant resupply of the acceptor NADP is mandatory. Several known stromal mechanisms aid in balancing the redox poise, but none of them utilizes the structurally highly similar coenzyme NAD(H). Using Arabidopsis (Arabidopsis thaliana) as a C3-model, we describe a pathway that employs the stromal enzyme PHOSPHOGLYCERATE DEHYDROGENASE 3 (PGDH3). We showed that PGDH3 exerts high NAD(H)-specificity and is active in photosynthesizing chloroplasts. PGDH3 withdrew its substrate 3-PGA directly from the CBBC. As a result, electrons become diverted from NADPH via the CBBC into the separate NADH redox pool. pgdh3 loss-of-function mutants revealed an overreduced NADP(H) redox pool but a more oxidized plastid NAD(H) pool compared to wild-type plants. As a result, photosystem I acceptor side limitation increased in pgdh3. Furthermore, pgdh3 plants displayed delayed CBBC activation, changes in nonphotochemical quenching, and altered proton motive force partitioning. Our fluctuating light-stress phenotyping data showed progressing photosystem II damage in pgdh3 mutants, emphasizing the significance of PGDH3 for plant performance under natural light environments. In summary, this study reveals an NAD(H)-specific mechanism in the stroma that aids in balancing the chloroplast redox poise. Consequently, the stromal NAD(H) pool may provide a promising target to manipulate plant photosynthesis.
在光合作用过程中,电子从受光激发的叶绿素分子沿着电子传递链传递到最终电子受体烟酰胺腺嘌呤二核苷酸磷酸(NADP),形成 NADPH,为卡尔文-本森-巴斯汉姆循环(CBBC)提供燃料。为了使光合作用反应完美进行,必须不断补充受体 NADP。几种已知的基质机制有助于平衡氧化还原平衡,但没有一种机制利用结构高度相似的辅酶 NAD(H)。我们使用拟南芥(Arabidopsis thaliana)作为 C3 模型,描述了一条利用基质酶磷酸甘油酸脱氢酶 3(PGDH3)的途径。我们表明,PGDH3 表现出高 NAD(H)特异性,并且在光合作用的叶绿体中具有活性。PGDH3 直接从 CBBC 中提取其底物 3-PGA。结果,电子通过 CBBC 从 NADPH 中分流,进入单独的 NADH 氧化还原池。pgdh3 功能丧失突变体显示 NADP(H)氧化还原池过度还原,但与野生型植物相比,质体 NAD(H)池更氧化。结果,PGDH3 增加了光系统 I 受体侧限制。此外,pgdh3 植物表现出 CBBC 激活延迟、非光化学猝灭变化和质子动力势分配改变。我们的波动光胁迫表型数据表明 pgdh3 突变体中光系统 II 损伤逐渐增加,强调了 PGDH3 在自然光照环境下对植物性能的重要性。总之,这项研究揭示了基质中一种有助于平衡叶绿体氧化还原平衡的 NAD(H)特异性机制。因此,基质 NAD(H)池可能为操纵植物光合作用提供一个有前途的目标。