Department of Physics, Pennsylvania State University, University Park, Pennsylvania, United States of America.
Eli and Edythe L. Broad Institute of MIT and Harvard, Cambridge, Massachusetts, United States of America.
PLoS Comput Biol. 2021 Mar 29;17(3):e1008690. doi: 10.1371/journal.pcbi.1008690. eCollection 2021 Mar.
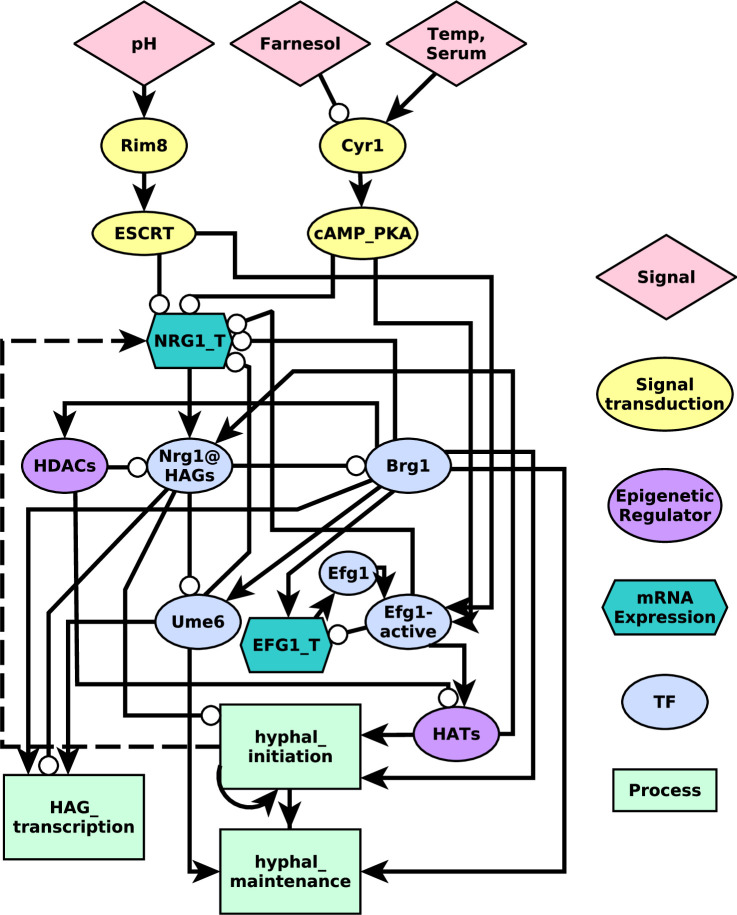
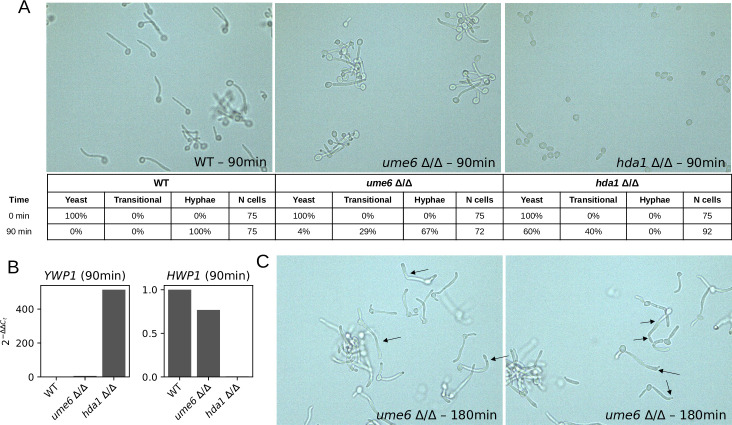
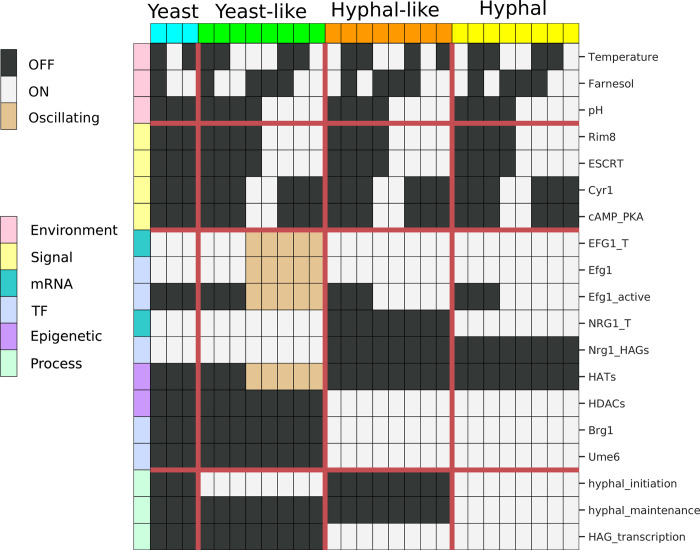
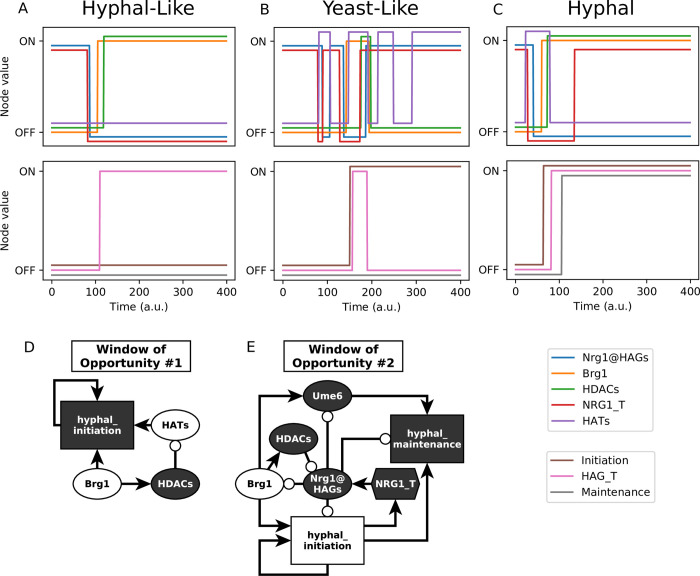
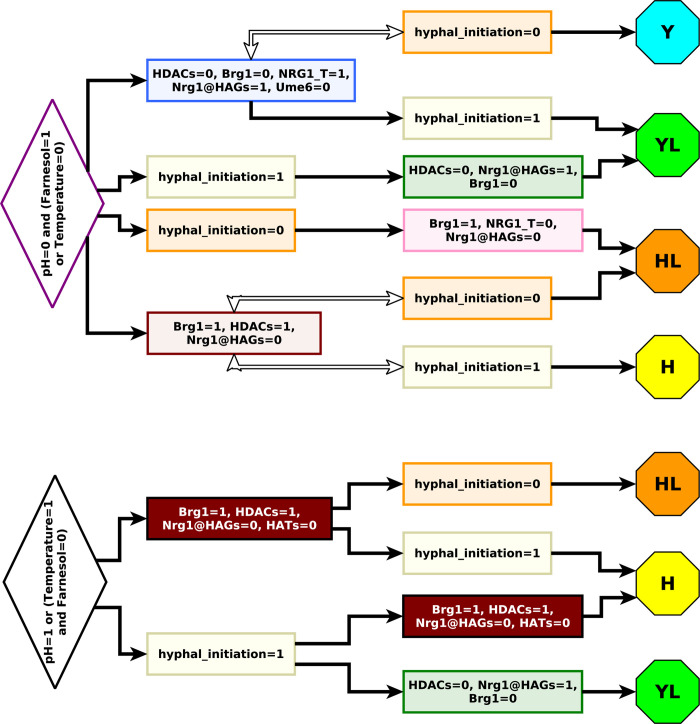
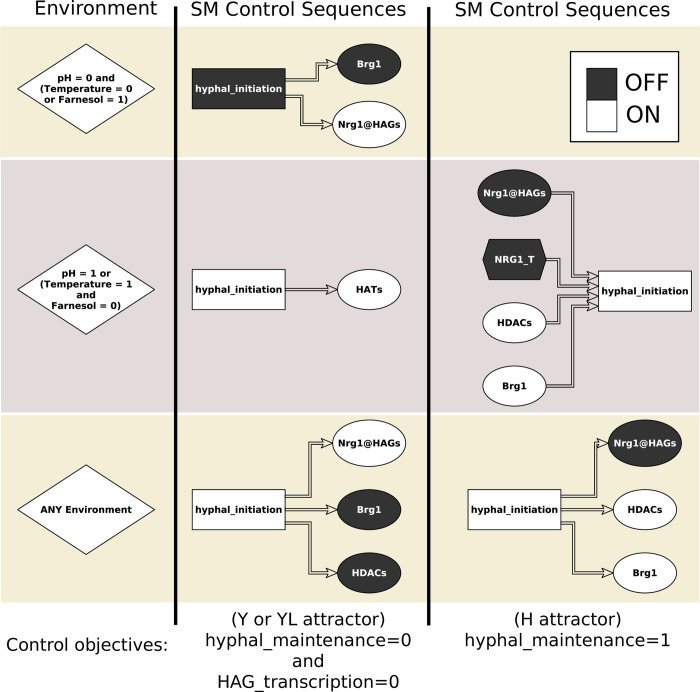
Candida albicans, an opportunistic fungal pathogen, is a significant cause of human infections, particularly in immunocompromised individuals. Phenotypic plasticity between two morphological phenotypes, yeast and hyphae, is a key mechanism by which C. albicans can thrive in many microenvironments and cause disease in the host. Understanding the decision points and key driver genes controlling this important transition and how these genes respond to different environmental signals is critical to understanding how C. albicans causes infections in the host. Here we build and analyze a Boolean dynamical model of the C. albicans yeast to hyphal transition, integrating multiple environmental factors and regulatory mechanisms. We validate the model by a systematic comparison to prior experiments, which led to agreement in 17 out of 22 cases. The discrepancies motivate alternative hypotheses that are testable by follow-up experiments. Analysis of this model revealed two time-constrained windows of opportunity that must be met for the complete transition from the yeast to hyphal phenotype, as well as control strategies that can robustly prevent this transition. We experimentally validate two of these control predictions in C. albicans strains lacking the transcription factor UME6 and the histone deacetylase HDA1, respectively. This model will serve as a strong base from which to develop a systems biology understanding of C. albicans morphogenesis.
白色念珠菌是一种机会性真菌病原体,是人类感染的重要原因,特别是在免疫功能低下的个体中。两种形态表型(酵母和菌丝)之间的表型可塑性是白色念珠菌在许多微环境中生存并在宿主中引起疾病的关键机制。了解控制这一重要转变的决策点和关键驱动基因,以及这些基因如何对不同的环境信号做出反应,对于理解白色念珠菌如何在宿主中引起感染至关重要。在这里,我们构建并分析了一个整合多种环境因素和调控机制的白色念珠菌酵母到菌丝体转变的布尔动态模型。我们通过与先前实验的系统比较来验证该模型,在 22 个案例中有 17 个案例达成了一致。差异促使提出了可通过后续实验验证的替代假设。对该模型的分析揭示了从酵母表型向菌丝表型完全转变必须满足的两个时间受限的机会窗口,以及可以稳健地阻止这种转变的控制策略。我们在分别缺乏转录因子 UME6 和组蛋白去乙酰化酶 HDA1 的白色念珠菌菌株中实验验证了这两个控制预测中的两个。这个模型将作为一个强大的基础,从中发展出对白色念珠菌形态发生的系统生物学理解。