Woods Kerry, Perry Carmen, Brühlmann Francis, Olias Philipp
Institute of Animal Pathology, Vetsuisse Faculty, University of Bern, Bern, Switzerland.
Front Cell Dev Biol. 2021 Apr 20;9:662805. doi: 10.3389/fcell.2021.662805. eCollection 2021.
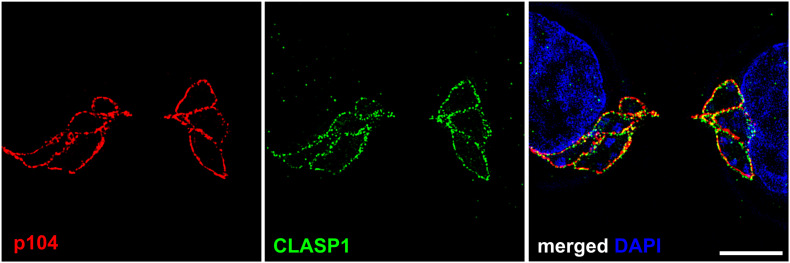
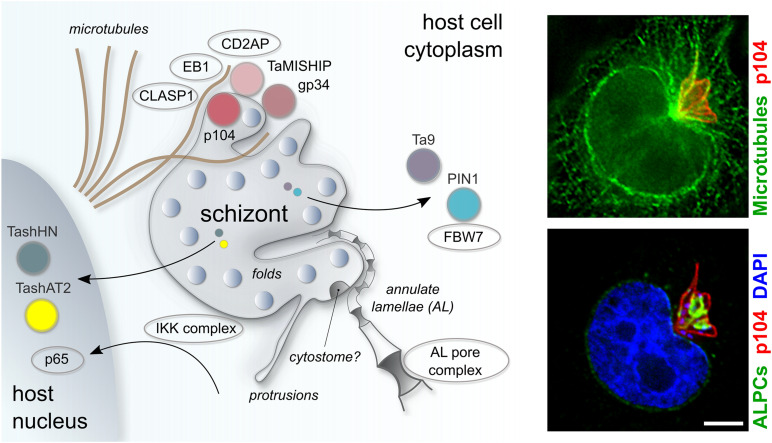
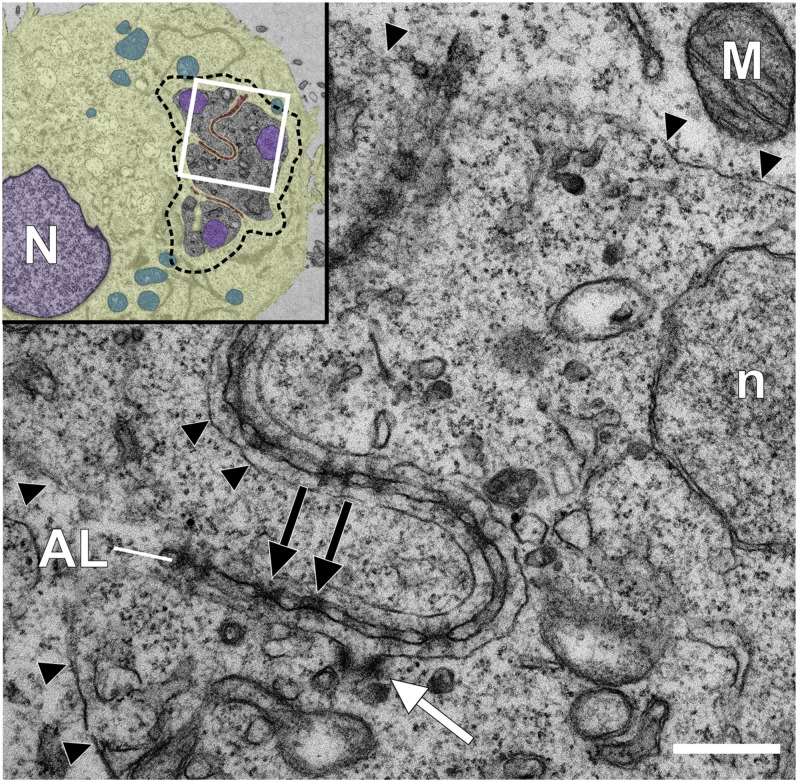
One of the first events that follows invasion of leukocytes by sporozoites is the destruction of the surrounding host cell membrane and the rapid association of the intracellular parasite with host microtubules. This is essential for the parasite to establish its niche within the cytoplasm of the invaded leukocyte and sets spp. apart from other members of the apicomplexan phylum such as and spp., which reside within the confines of a host-derived parasitophorous vacuole. After establishing infection, transforming species (, ) significantly rewire the signaling pathways of their bovine host cell, causing continual proliferation and resistance to ligand-induced apoptosis, and conferring invasive properties on the parasitized cell. Having transformed its target cell, hijacks the mitotic machinery to ensure its persistence in the cytoplasm of the dividing cell. Some of the parasite and bovine proteins involved in parasite-microtubule interactions have been fairly well characterized, and the schizont expresses at least two proteins on its membrane that contain conserved microtubule binding motifs. -encoded proteins have been shown to be translocated to the host cell cytoplasm and nucleus where they have the potential to directly modify signaling pathways and host gene expression. However, little is known about their mode of action, and even less about how these proteins are secreted by the parasite and trafficked to their target location. In this review we explore the strategies employed by to transform leukocytes, from sporozoite invasion until immortalization of the host cell has been established. We discuss the recent description of nuclear pore-like complexes that accumulate on membranes close to the schizont surface. Finally, we consider putative mechanisms of protein and nutrient exchange that might occur between the parasite and the host. We focus in particular on differences and similarities with recent discoveries in and species.
子孢子侵入白细胞后发生的首批事件之一是周围宿主细胞膜的破坏以及细胞内寄生虫与宿主微管的快速结合。这对于寄生虫在被侵入白细胞的细胞质中建立其生态位至关重要,并且使该物种与顶复门的其他成员(如疟原虫属和巴贝斯虫属)区分开来,后者寄生于宿主来源的寄生泡内。在建立感染后,转化型泰勒虫属物种(环形泰勒虫、瑟氏泰勒虫)会显著重塑其牛宿主细胞的信号通路,导致细胞持续增殖并抵抗配体诱导的凋亡,并赋予被寄生细胞侵袭特性。在转化其靶细胞后,环形泰勒虫会劫持有丝分裂机制以确保其在分裂细胞的细胞质中持续存在。一些参与寄生虫与微管相互作用的寄生虫和牛蛋白质已得到相当充分的表征,并且裂殖体在其膜上表达至少两种含有保守微管结合基序的蛋白质。泰勒虫编码的蛋白质已被证明可转运至宿主细胞的细胞质和细胞核,在那里它们有可能直接改变信号通路和宿主基因表达。然而,人们对它们的作用方式知之甚少,对于这些蛋白质如何由寄生虫分泌并运输到其靶位点更是了解甚少。在本综述中,我们探讨了泰勒虫从子孢子侵入到宿主细胞永生化建立过程中用于转化白细胞的策略。我们讨论了最近对在裂殖体表面附近的膜上积累的核孔样复合体的描述。最后,我们考虑了寄生虫与宿主之间可能发生的蛋白质和营养物质交换的推定机制。我们特别关注与疟原虫属和巴贝斯虫属物种最近发现的异同。