Neurobiology Division, MRC Laboratory of Molecular Biology, Cambridge, UK.
IST Austria, Klosterneuburg, Austria.
Nature. 2021 Jun;594(7863):454-458. doi: 10.1038/s41586-021-03613-0. Epub 2021 Jun 2.
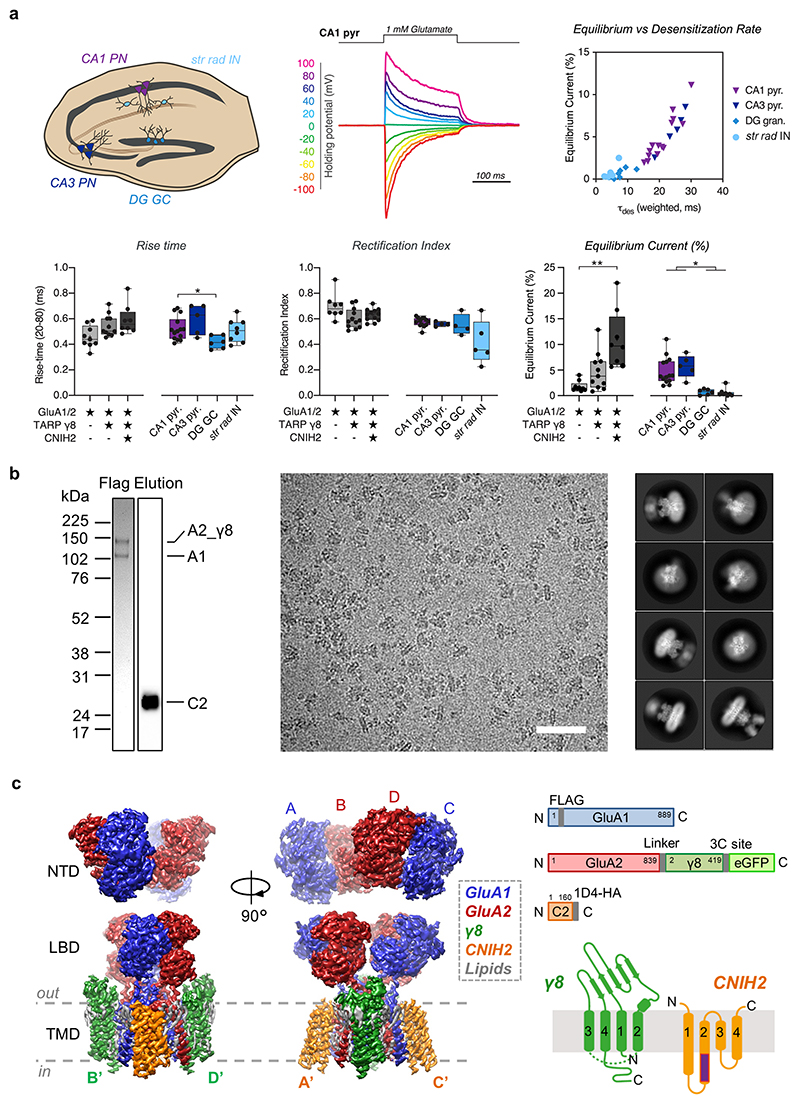
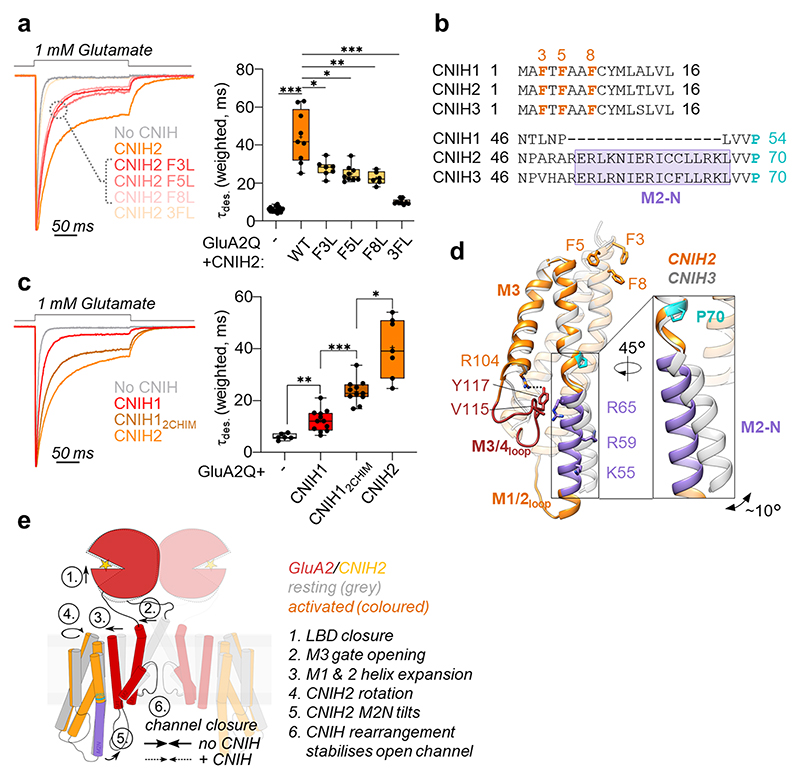
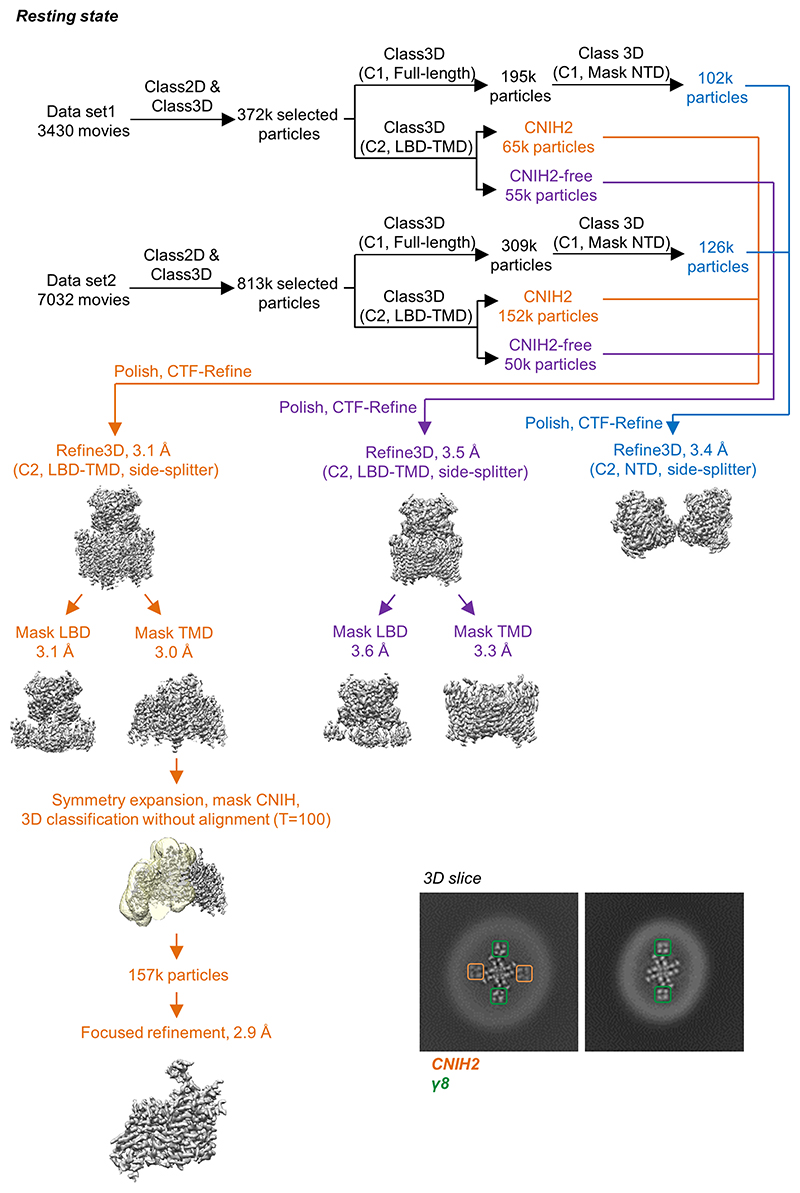
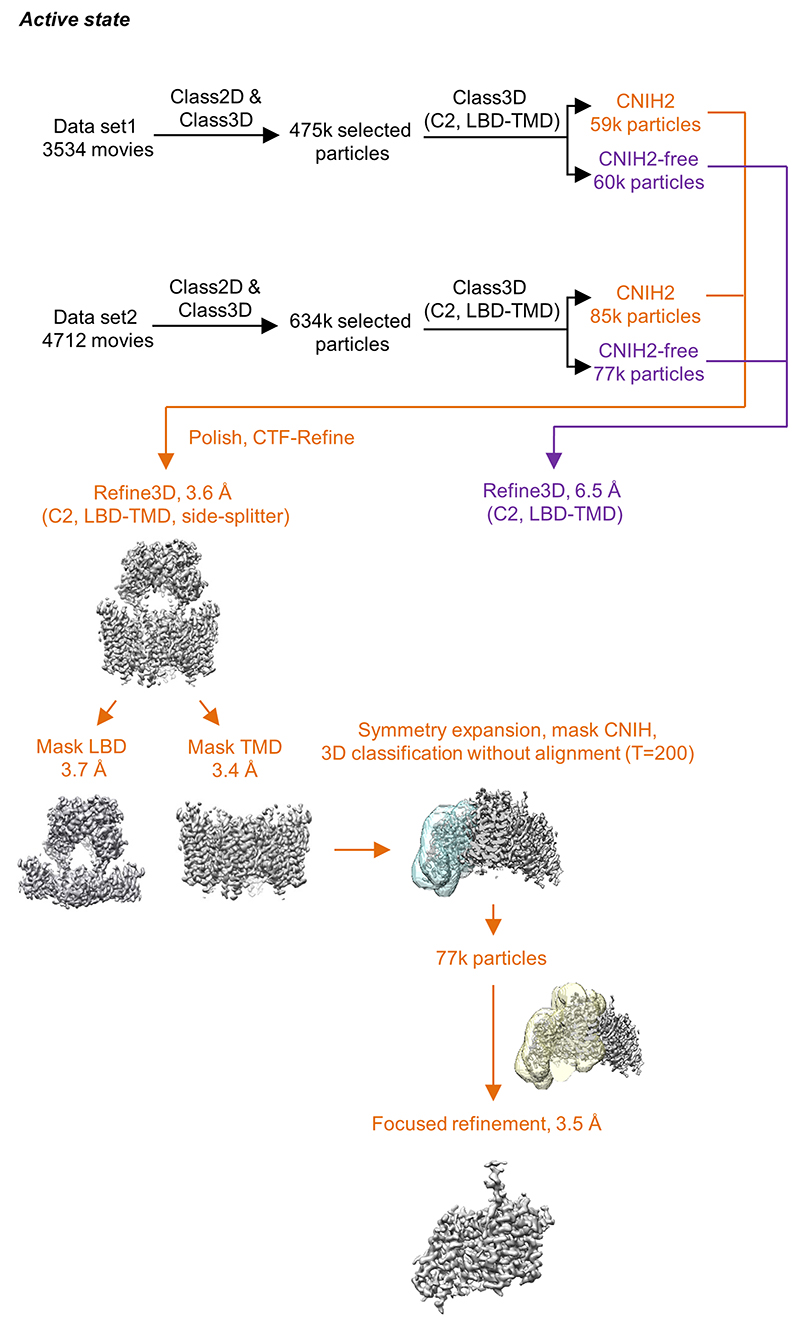
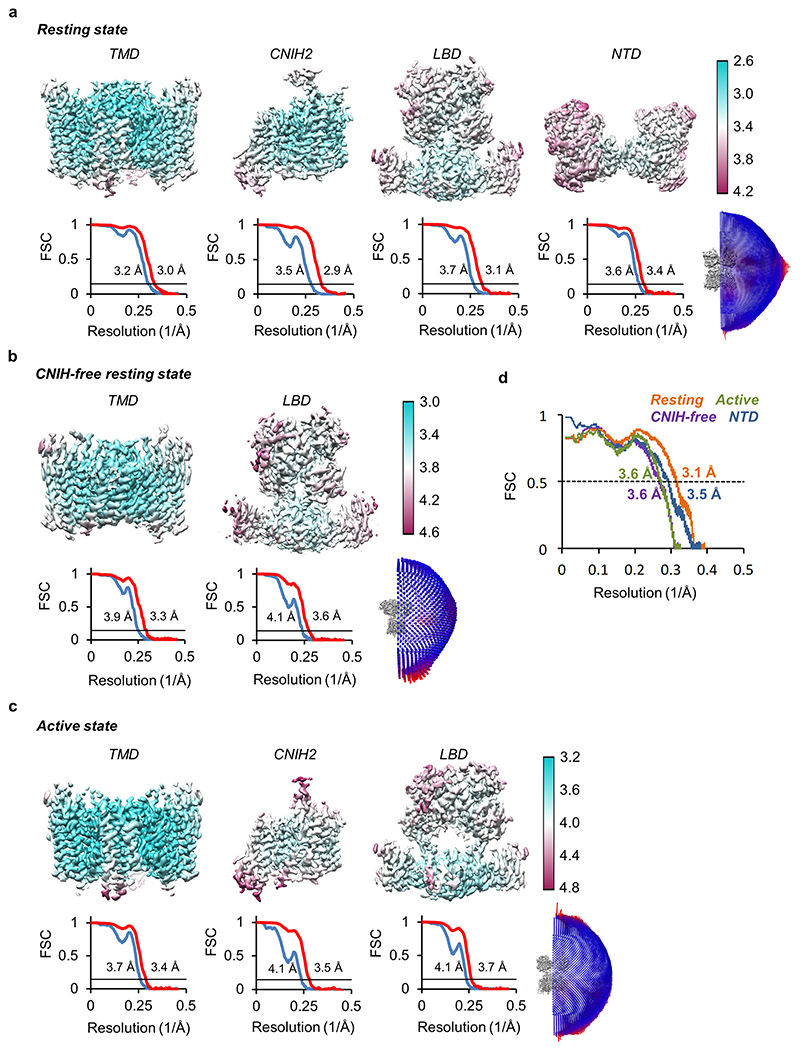
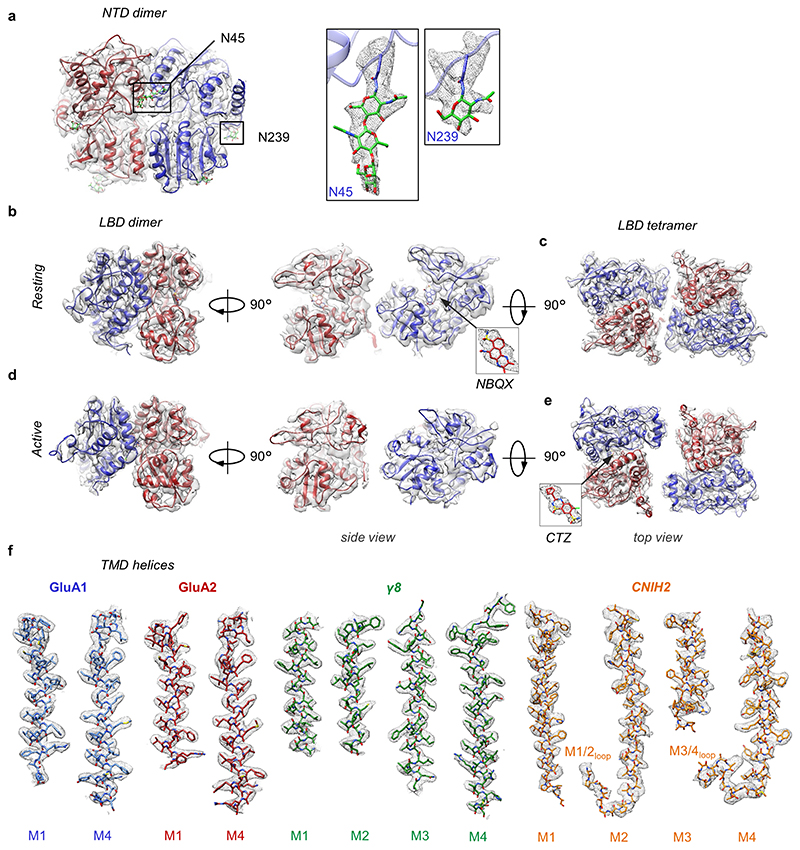
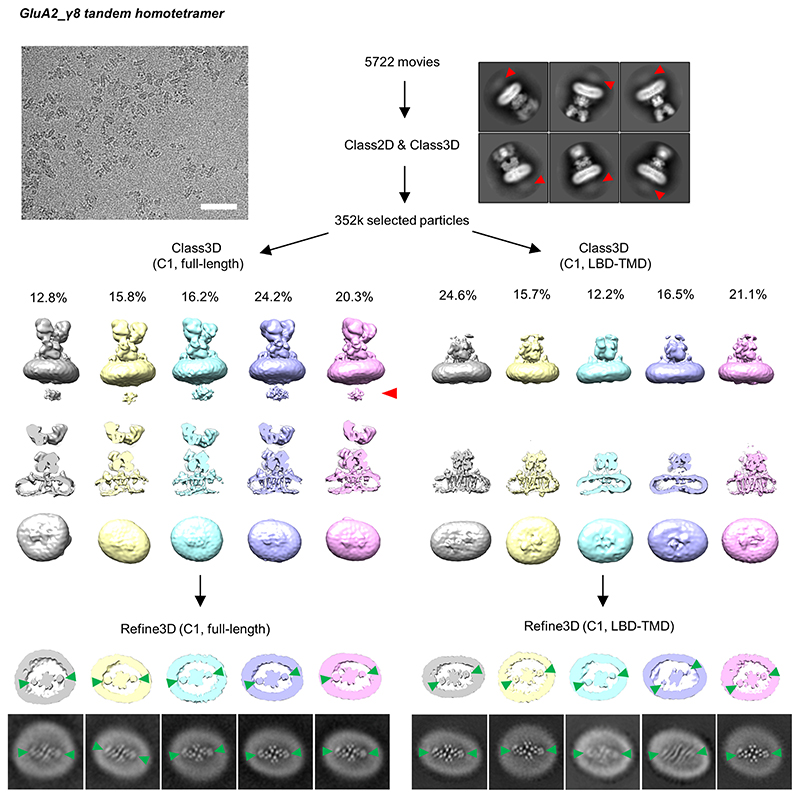
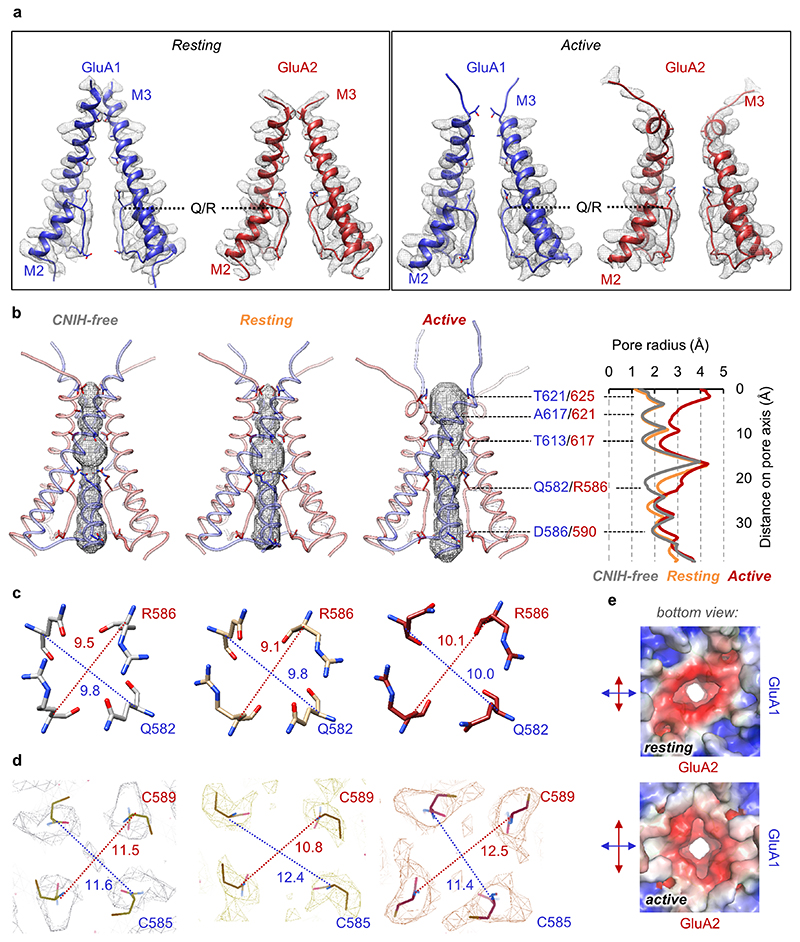
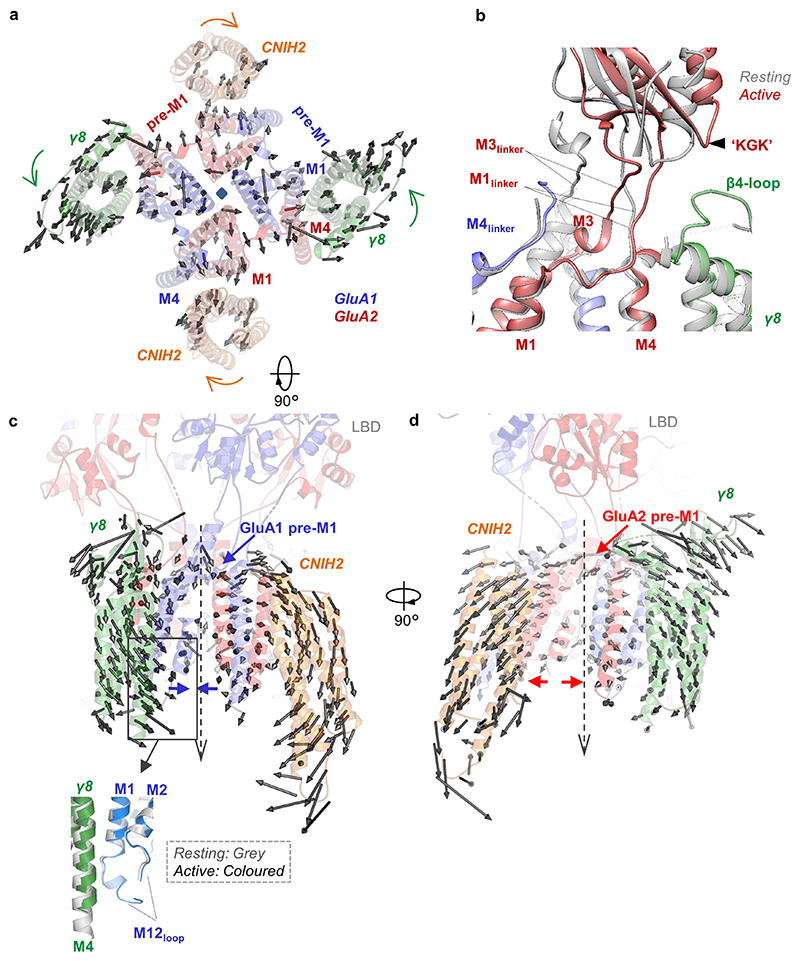
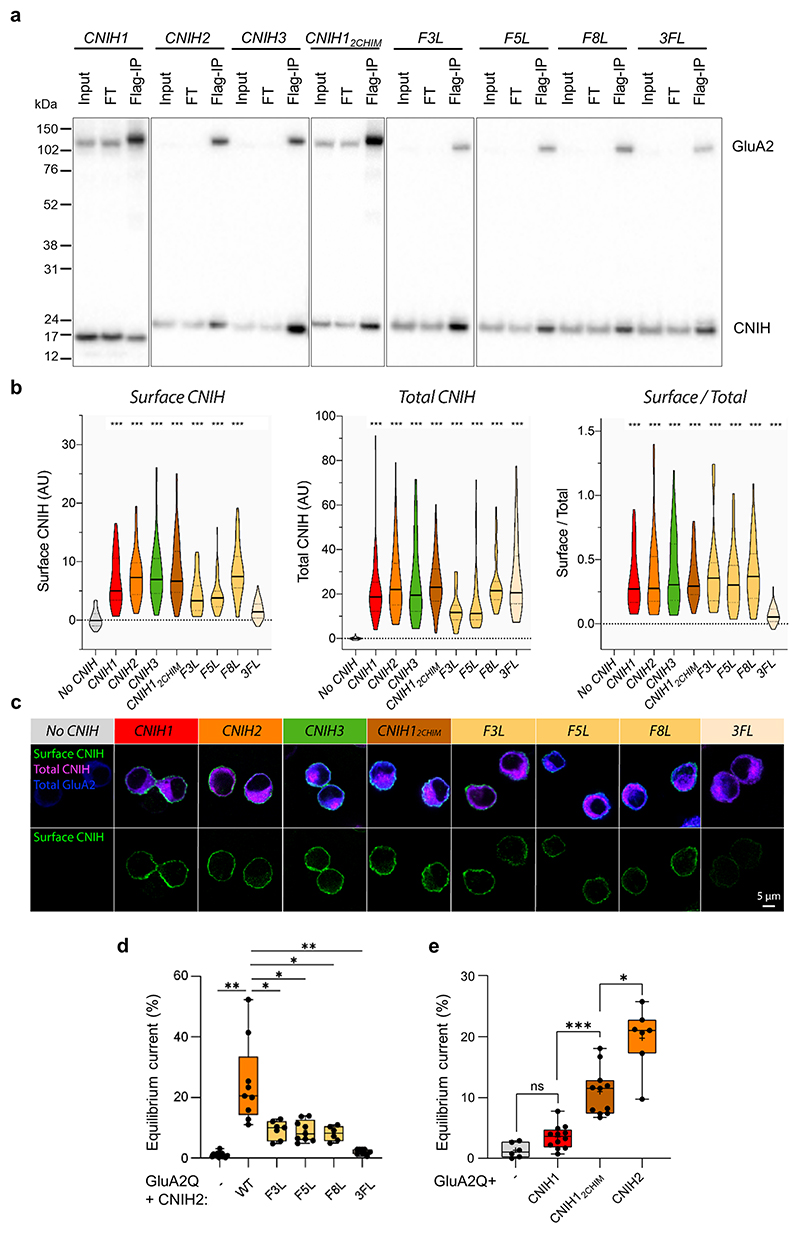
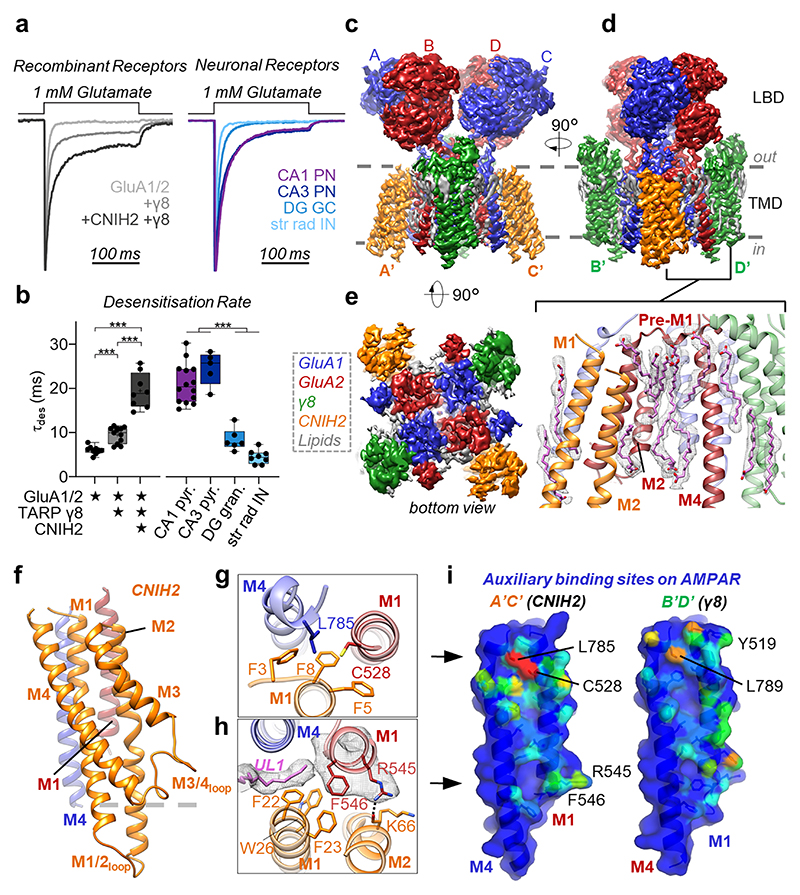
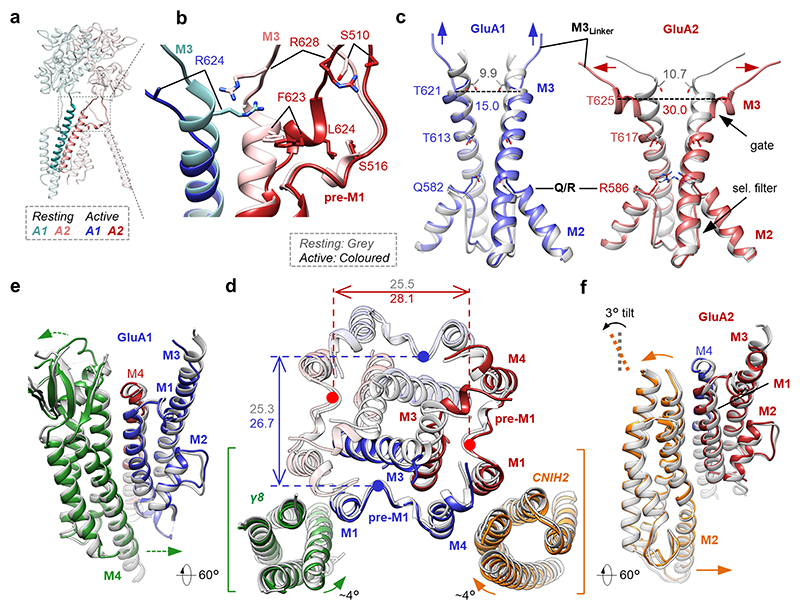
AMPA receptors (AMPARs) mediate the majority of excitatory transmission in the brain and enable the synaptic plasticity that underlies learning. A diverse array of AMPAR signalling complexes are established by receptor auxiliary subunits, which associate with the AMPAR in various combinations to modulate trafficking, gating and synaptic strength. However, their mechanisms of action are poorly understood. Here we determine cryo-electron microscopy structures of the heteromeric GluA1-GluA2 receptor assembled with both TARP-γ8 and CNIH2, the predominant AMPAR complex in the forebrain, in both resting and active states. Two TARP-γ8 and two CNIH2 subunits insert at distinct sites beneath the ligand-binding domains of the receptor, with site-specific lipids shaping each interaction and affecting the gating regulation of the AMPARs. Activation of the receptor leads to asymmetry between GluA1 and GluA2 along the ion conduction path and an outward expansion of the channel triggers counter-rotations of both auxiliary subunit pairs, promoting the active-state conformation. In addition, both TARP-γ8 and CNIH2 pivot towards the pore exit upon activation, extending their reach for cytoplasmic receptor elements. CNIH2 achieves this through its uniquely extended M2 helix, which has transformed this endoplasmic reticulum-export factor into a powerful AMPAR modulator that is capable of providing hippocampal pyramidal neurons with their integrative synaptic properties.
AMPA 受体 (AMPARs) 介导大脑中大多数兴奋性传递,并实现了支持学习的突触可塑性。受体辅助亚基建立了多种多样的 AMPAR 信号复合物,它们与 AMPAR 以各种组合结合,从而调节转运、门控和突触强度。然而,它们的作用机制仍不清楚。在这里,我们确定了异源 GluA1-GluA2 受体与 TARP-γ8 和 CNIH2 组装的冷冻电镜结构,这两种复合物是前脑的主要 AMPAR 复合物,处于静息和激活状态。两个 TARP-γ8 和两个 CNIH2 亚基插入受体配体结合域下方的独特位点,具有特定于位点的脂质,形成每种相互作用,并影响 AMPAR 的门控调节。受体的激活导致沿离子传导路径在 GluA1 和 GluA2 之间产生不对称性,并且通道的向外扩展触发两个辅助亚基对的反向旋转,促进了激活状态构象。此外,在激活时,TARP-γ8 和 CNIH2 都向孔出口枢转,延伸其对细胞质受体元件的作用范围。CNIH2 通过其独特延伸的 M2 螺旋实现这一点,这将这种内质网出口因子转化为一种强大的 AMPAR 调节剂,能够为海马锥体神经元提供其整合的突触特性。