Program of Chemical Biology, University of Michigan, Ann Arbor, Michigan, USA; Life Sciences Institute, University of Michigan, Ann Arbor, Michigan, USA.
Department of Medicine, Division of Hematology and Oncology, Vanderbilt University Medical Center, Nashville, Tennessee, USA; Department of Genetics, Vanderbilt University, Nashville, Tennessee, USA.
J Biol Chem. 2021 Jul;297(1):100898. doi: 10.1016/j.jbc.2021.100898. Epub 2021 Jun 19.
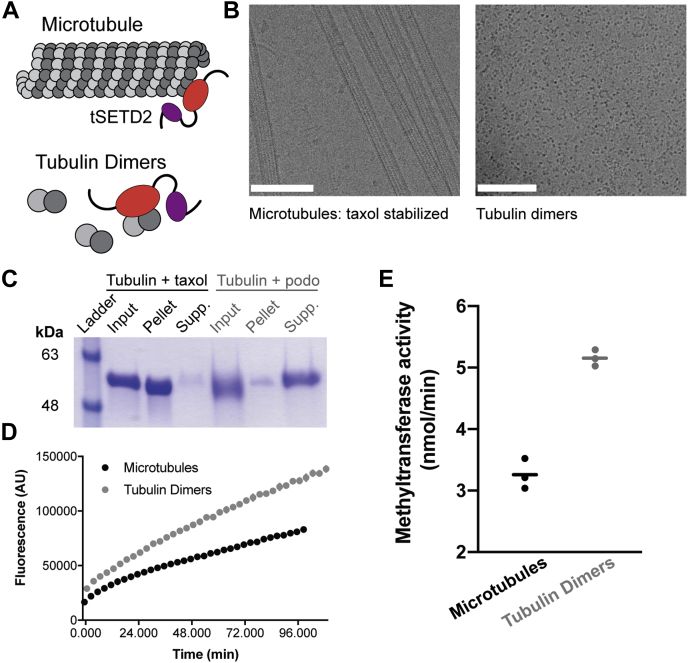
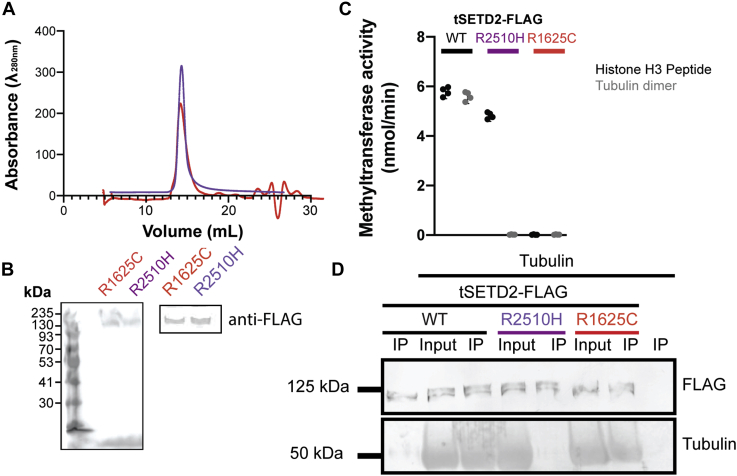
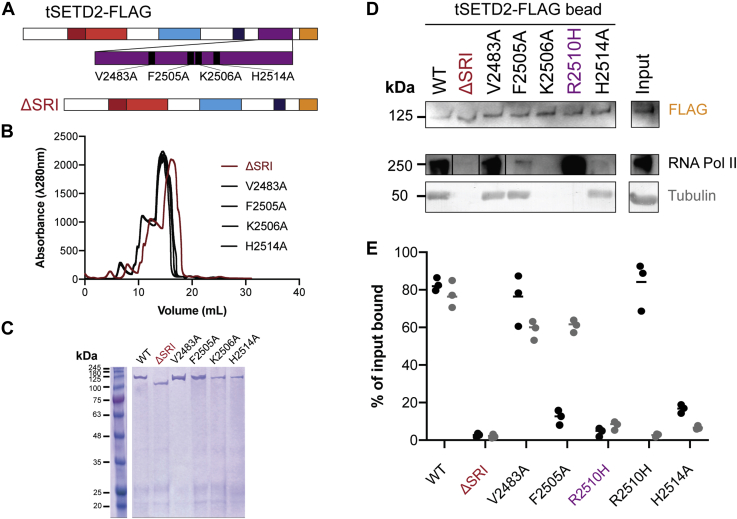
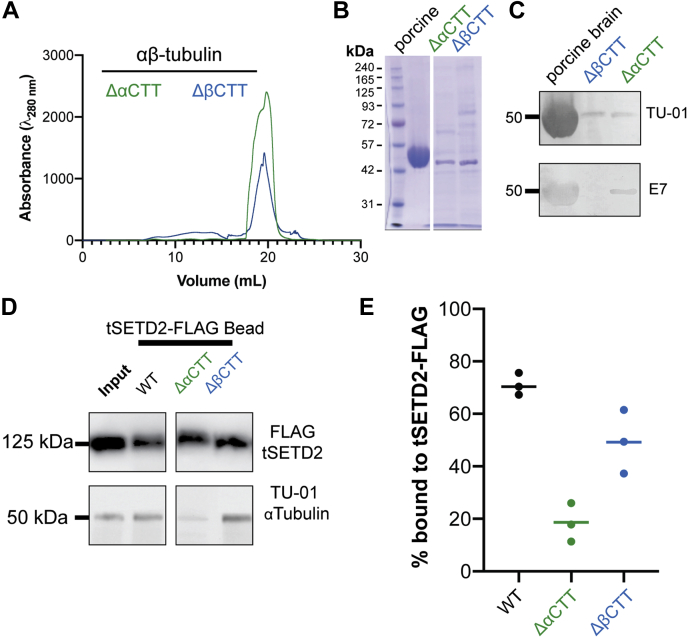
Post-translational modifications to tubulin are important for many microtubule-based functions inside cells. It was recently shown that methylation of tubulin by the histone methyltransferase SETD2 occurs on mitotic spindle microtubules during cell division, with its absence resulting in mitotic defects. However, the catalytic mechanism of methyl addition to tubulin is unclear. We used a truncated version of human wild type SETD2 (tSETD2) containing the catalytic SET and C-terminal Set2-Rpb1-interacting (SRI) domains to investigate the biochemical mechanism of tubulin methylation. We found that recombinant tSETD2 had a higher activity toward tubulin dimers than polymerized microtubules. Using recombinant single-isotype tubulin, we demonstrated that methylation was restricted to lysine 40 of α-tubulin. We then introduced pathogenic mutations into tSETD2 to probe the recognition of histone and tubulin substrates. A mutation in the catalytic domain (R1625C) allowed tSETD2 to bind to tubulin but not methylate it, whereas a mutation in the SRI domain (R2510H) caused loss of both tubulin binding and methylation. Further investigation of the role of the SRI domain in substrate binding found that mutations within this region had differential effects on the ability of tSETD2 to bind to tubulin versus the binding partner RNA polymerase II for methylating histones in vivo, suggesting distinct mechanisms for tubulin and histone methylation by SETD2. Finally, we found that substrate recognition also requires the negatively charged C-terminal tail of α-tubulin. Together, this study provides a framework for understanding how SETD2 serves as a dual methyltransferase for both histone and tubulin methylation.
微管蛋白的翻译后修饰对于细胞内许多基于微管的功能至关重要。最近有研究表明,组蛋白甲基转移酶SETD2对微管蛋白的甲基化发生在细胞分裂期间的有丝分裂纺锤体微管上,缺乏这种甲基化会导致有丝分裂缺陷。然而,向微管蛋白添加甲基的催化机制尚不清楚。我们使用了包含催化性SET和C末端Set2-Rpb1相互作用(SRI)结构域的人类野生型SETD2截短版本(tSETD2)来研究微管蛋白甲基化的生化机制。我们发现重组tSETD2对微管蛋白二聚体的活性高于聚合微管。使用重组单亚型微管蛋白,我们证明甲基化仅限于α-微管蛋白的赖氨酸40。然后,我们在tSETD2中引入致病突变以探究对组蛋白和微管蛋白底物的识别。催化结构域中的一个突变(R1625C)使tSETD2能够结合微管蛋白但不能使其甲基化,而SRI结构域中的一个突变(R2510H)导致微管蛋白结合和甲基化均丧失。对SRI结构域在底物结合中作用的进一步研究发现,该区域内的突变对tSETD2在体内结合微管蛋白与结合伴侣RNA聚合酶II以甲基化组蛋白的能力有不同影响,这表明SETD2对微管蛋白和组蛋白甲基化的机制不同。最后,我们发现底物识别还需要α-微管蛋白带负电荷的C末端尾巴。总之,这项研究为理解SETD2如何作为组蛋白和微管蛋白甲基化的双重甲基转移酶提供了一个框架。