Department of Physiology & Neuroscience, Zilkha Neurogenetic Institute, Keck School of Medicine, University of Southern California, Los Angeles, CA, USA.
Department of Emergency Medicine, Regions Hospital, St. Paul, MN, USA.
Nat Commun. 2021 Jul 13;12(1):4272. doi: 10.1038/s41467-021-24411-2.
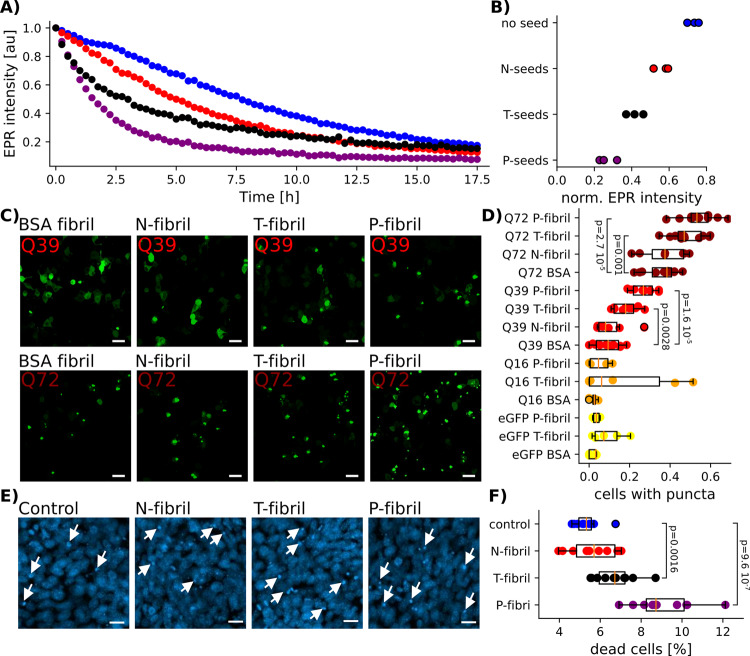
The first exon of the huntingtin protein (HTTex1) important in Huntington's disease (HD) can form cross-β fibrils of varying toxicity. We find that the difference between these fibrils is the degree of entanglement and dynamics of the C-terminal proline-rich domain (PRD) in a mechanism analogous to polyproline film formation. In contrast to fibril strains found for other cross-β fibrils, these HTTex1 fibril types can be interconverted. This is because the structure of their polyQ fibril core remains unchanged. Further, we find that more toxic fibrils of low entanglement have higher affinities for protein interactors and are more effective seeds for recombinant HTTex1 and HTTex1 in cells. Together these data show how the structure of a framing sequence at the surface of a fibril can modulate seeding, protein-protein interactions, and thereby toxicity in neurodegenerative disease.
亨廷顿病(HD)中重要的亨廷顿蛋白(HTTex1)的第一个外显子可以形成具有不同毒性的交叉β纤维。我们发现,这些纤维之间的差异在于 C 端脯氨酸丰富结构域(PRD)的缠结程度和动力学,这一机制类似于多聚脯氨酸膜的形成。与其他交叉β纤维的纤维菌株不同,这些 HTTex1 纤维类型可以相互转换。这是因为它们的多聚 Q 纤维核心结构保持不变。此外,我们发现缠结程度较低的毒性纤维具有更高的蛋白相互作用亲和力,并且对于重组 HTTex1 和 HTTex1 在细胞中的成核作用更有效。这些数据共同表明,纤维表面框架序列的结构如何调节成核、蛋白-蛋白相互作用,从而影响神经退行性疾病的毒性。