Department of Zoology, University of Cambridge, Downing St., Cambridge, United Kingdom.
Smithsonian Tropical Research Institute, Gamboa, Panama.
Elife. 2021 Jul 19;10:e68549. doi: 10.7554/eLife.68549.
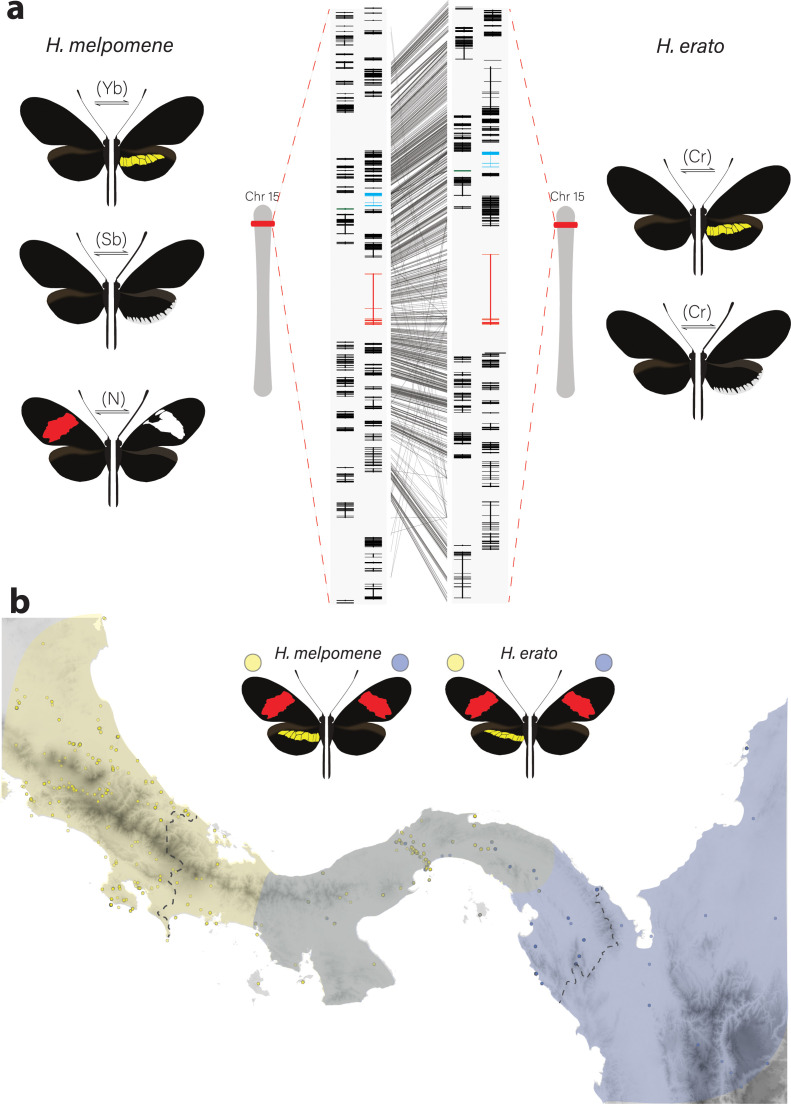
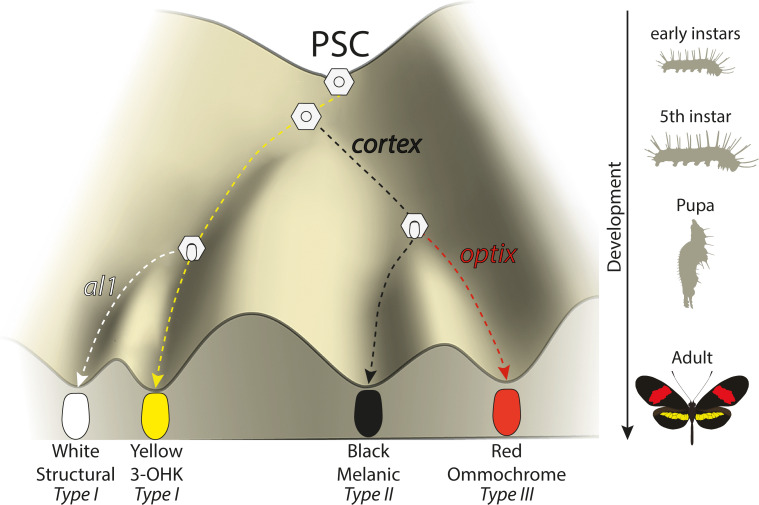
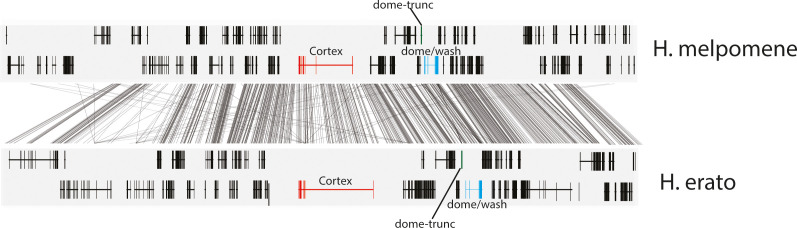
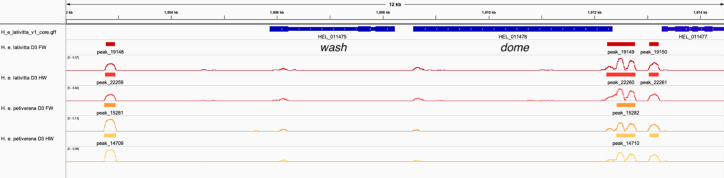
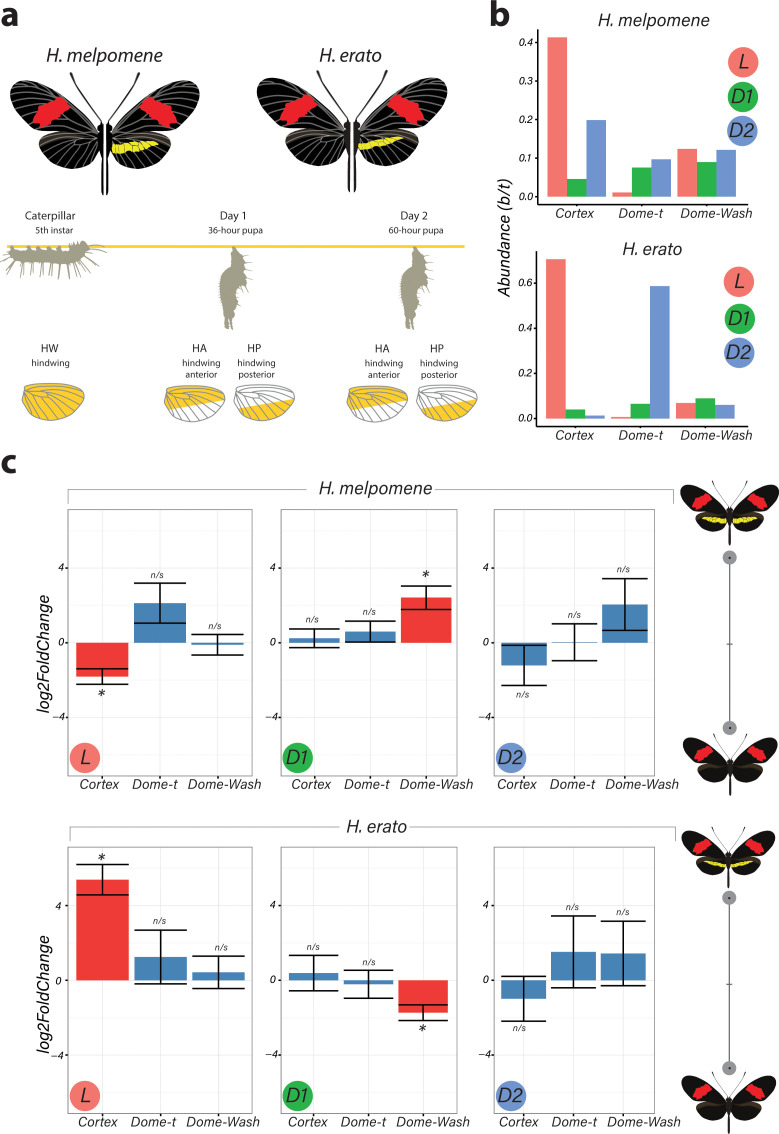
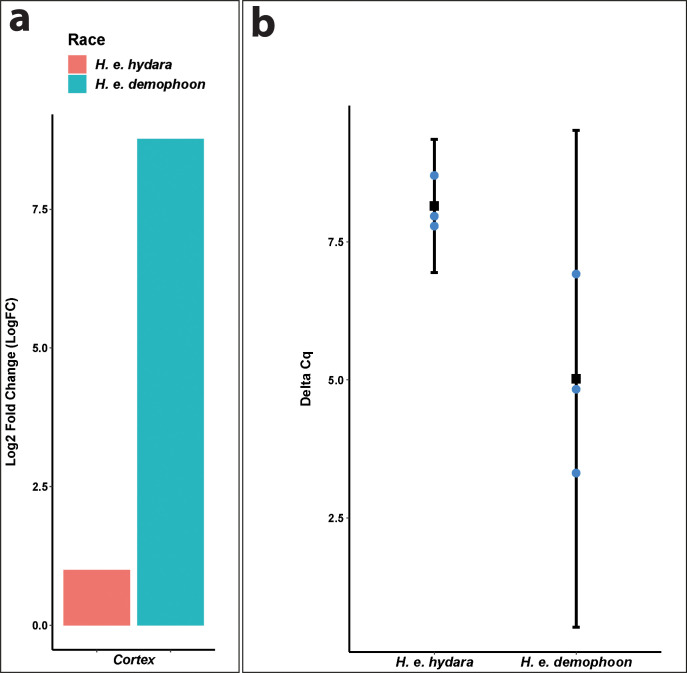
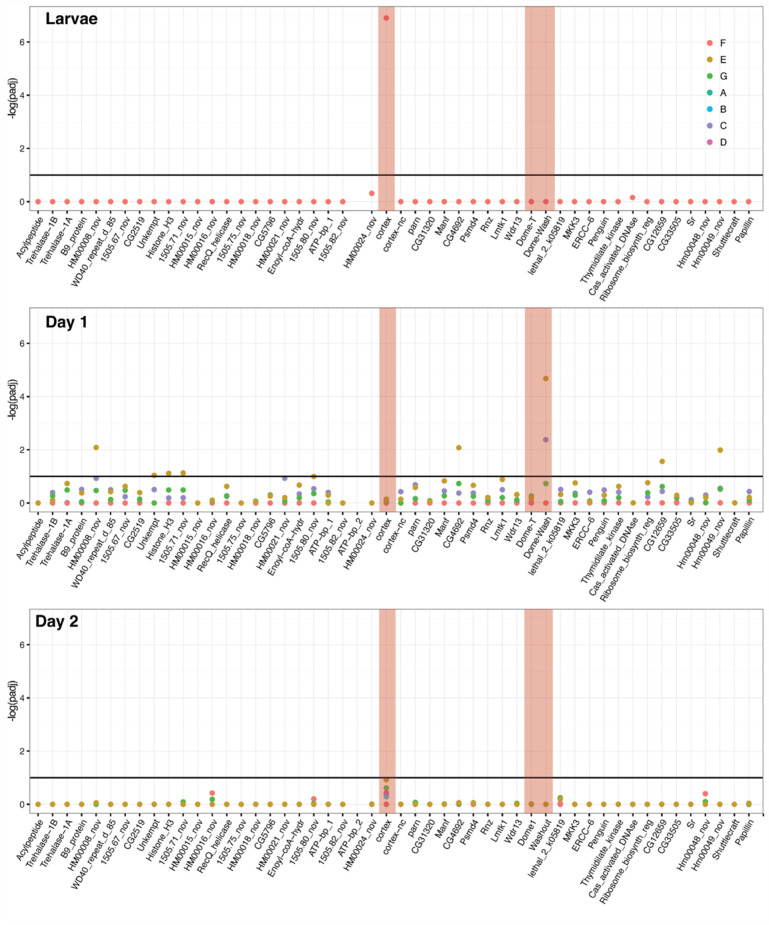
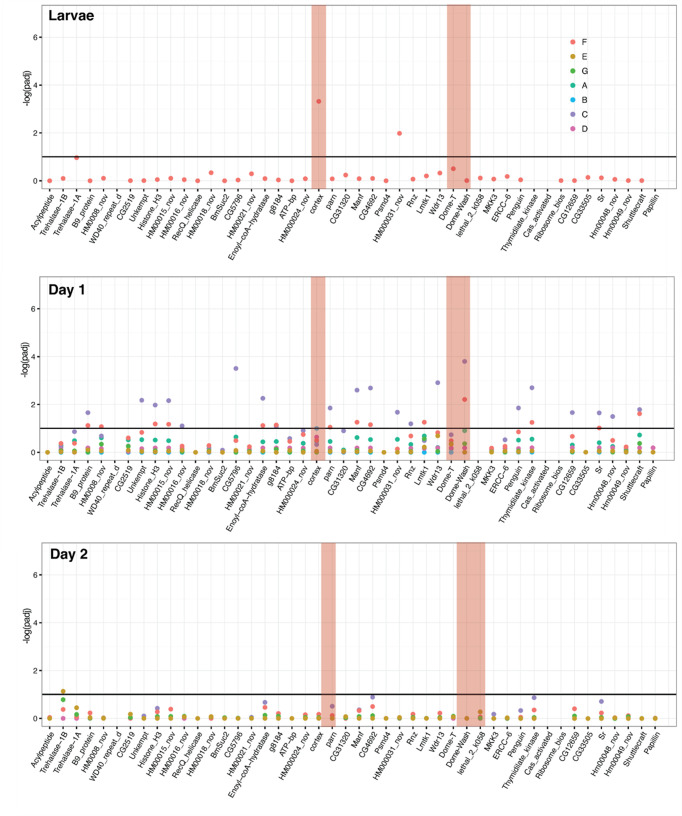
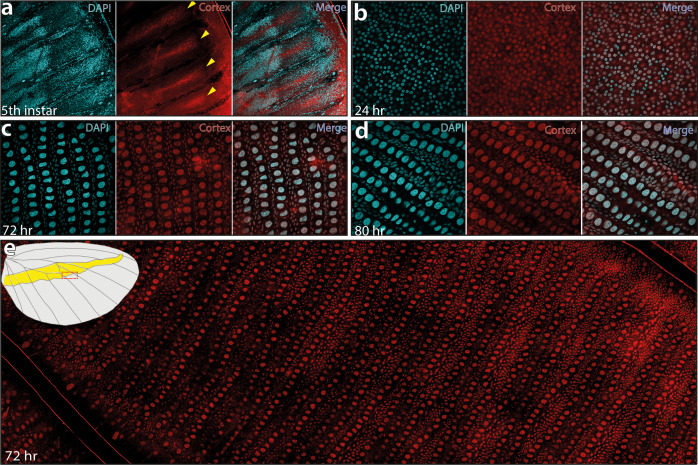
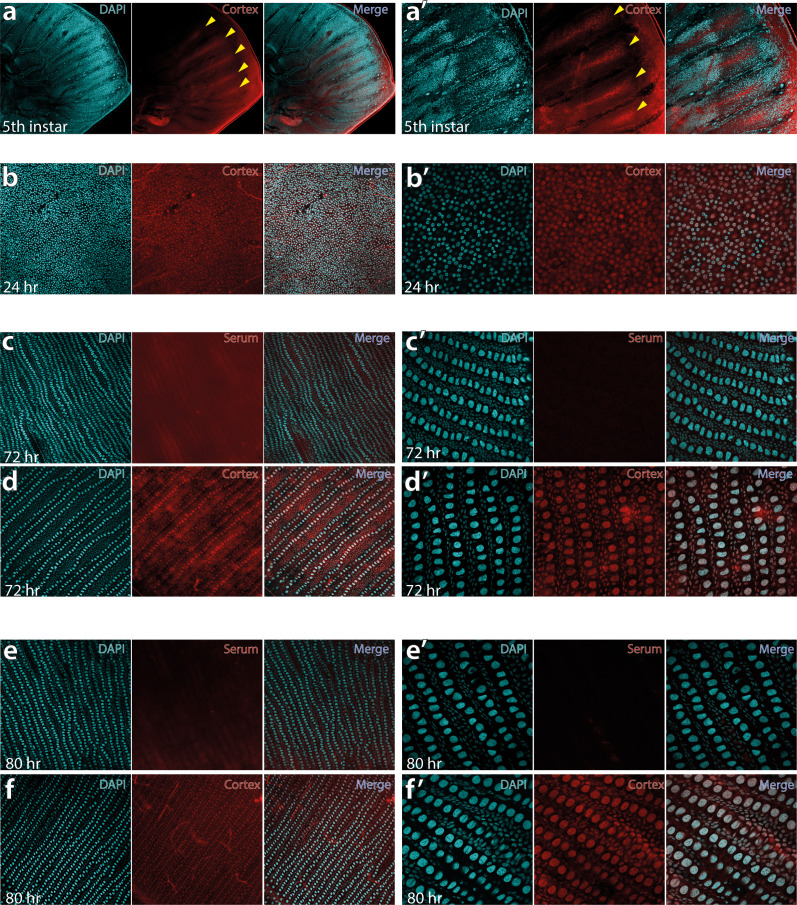
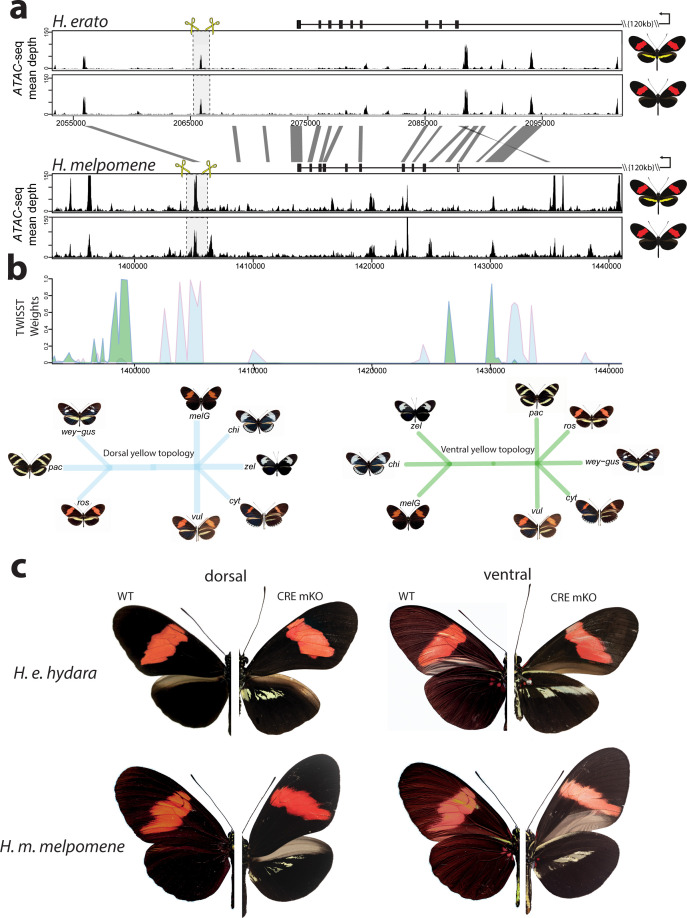
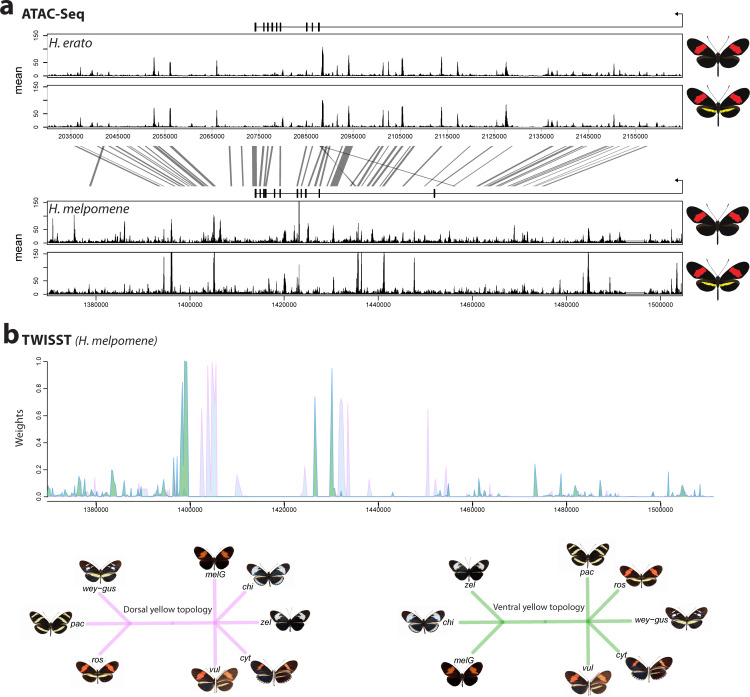
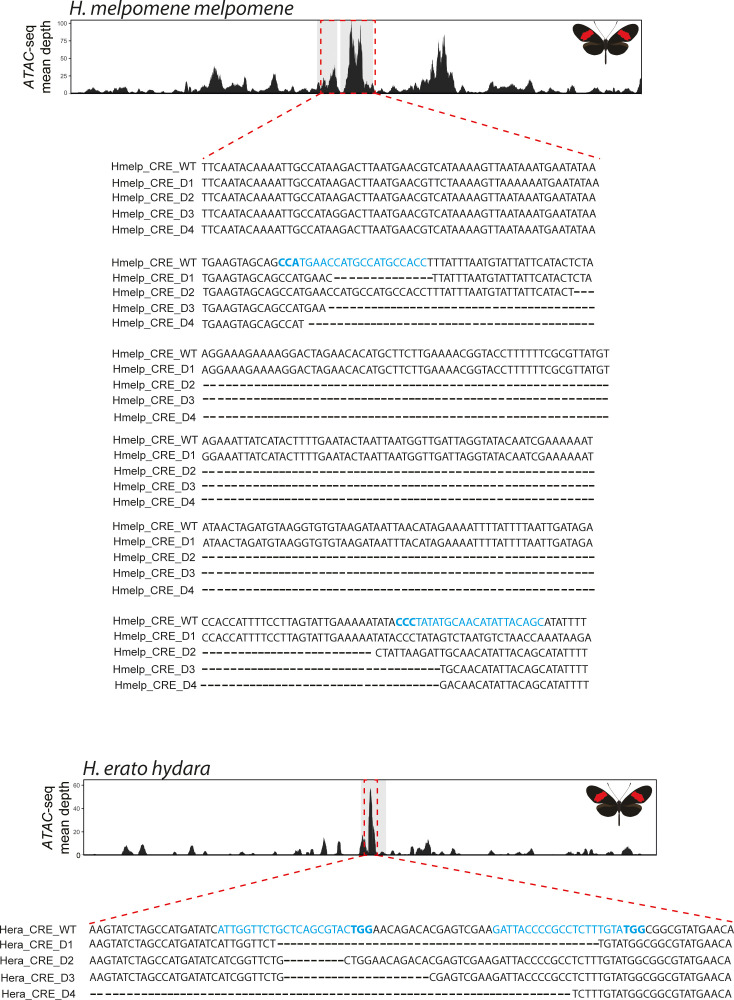
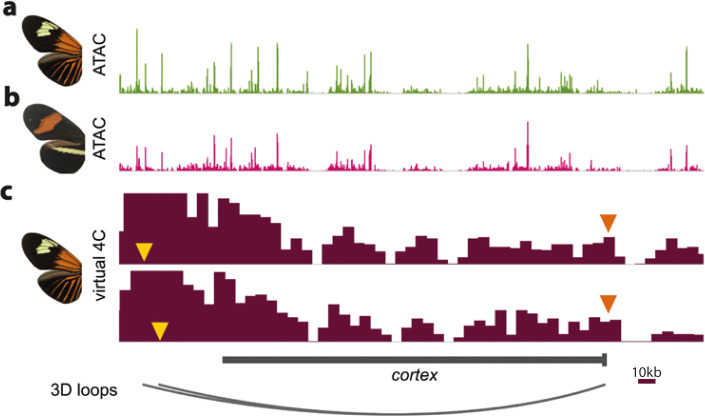
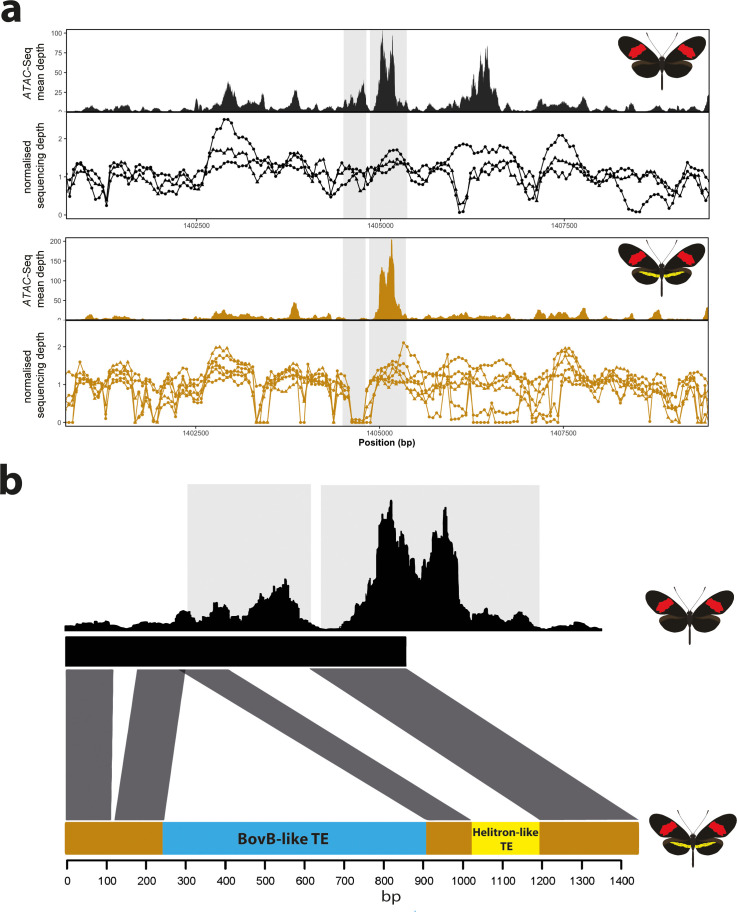
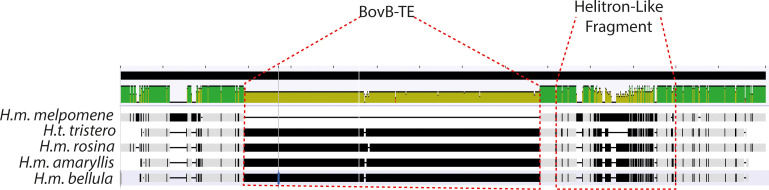
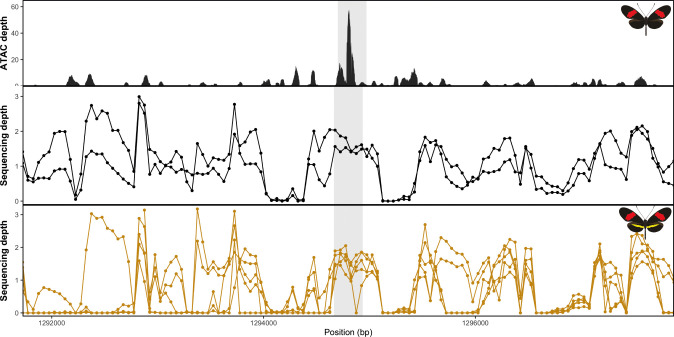
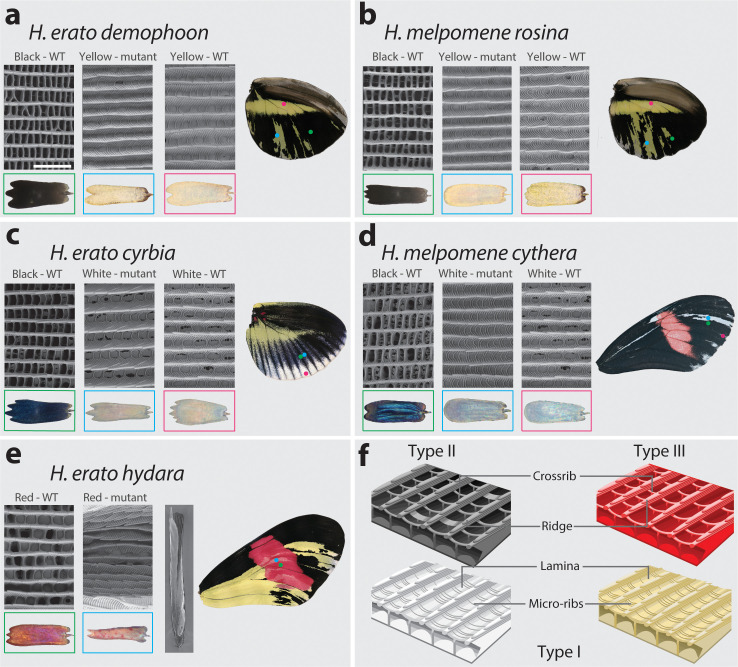
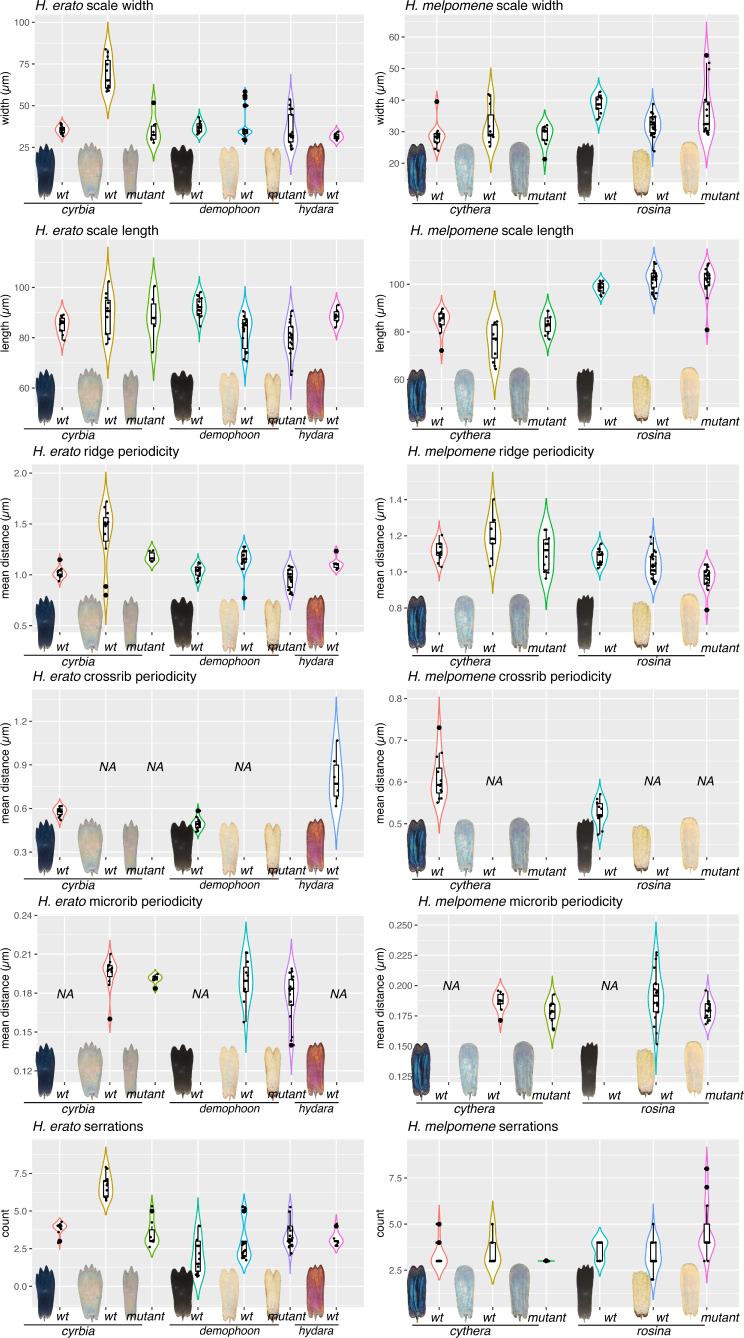
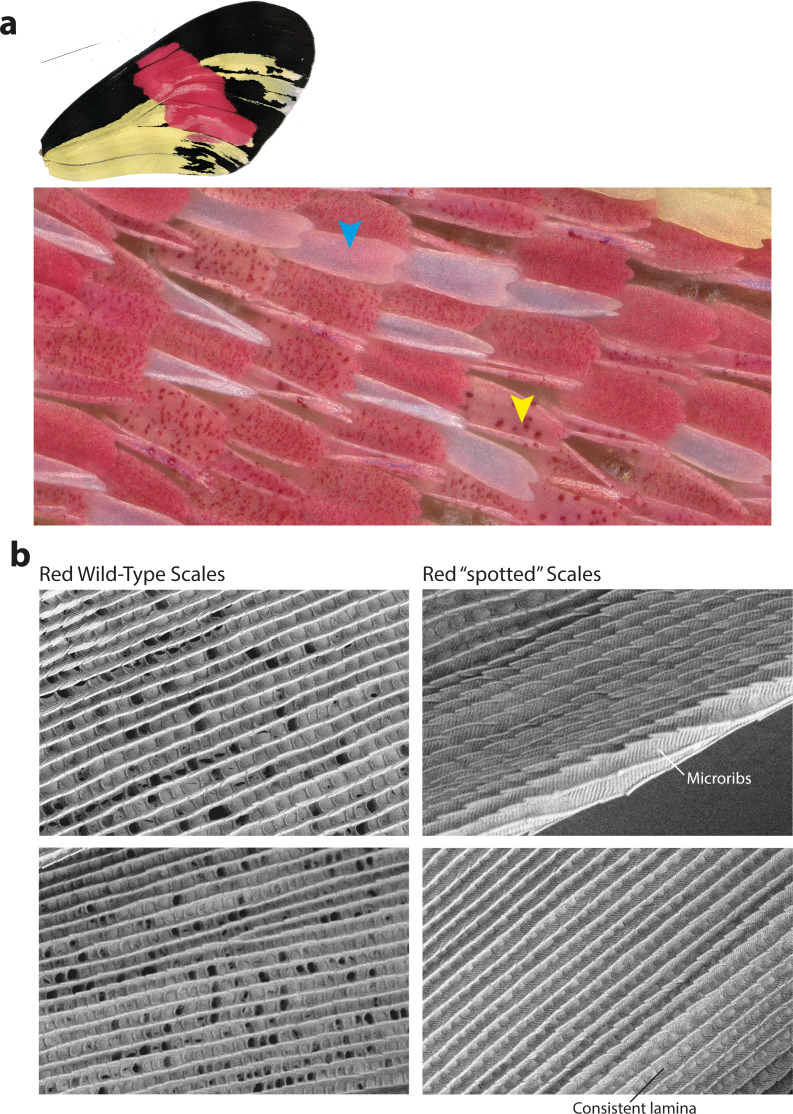
In butterflies, wing colour pattern diversity and scale types are controlled by a few genes of large effect that regulate colour pattern switches between morphs and species across a large mimetic radiation. One of these genes, , has been repeatedly associated with colour pattern evolution in butterflies. Here we carried out CRISPR knockouts in multiple species and show that is a major determinant of scale cell identity. Chromatin accessibility profiling and introgression scans identified -regulatory regions associated with discrete phenotypic switches. CRISPR perturbation of these regions in black hindwing genotypes recreated a yellow bar, revealing their spatially limited activity. In the lineage, the candidate CRE from yellow-barred phenotype morphs is interrupted by a transposable element, suggesting that -regulatory structural variation underlies these mimetic adaptations. Our work shows that functionally controls scale colour fate and that its -regulatory regions control a phenotypic switch in a modular and pattern-specific fashion.
在蝴蝶中,翅膀颜色图案的多样性和鳞片类型由少数几个大效应基因控制,这些基因调节着不同形态和物种之间的颜色图案转换,形成了一个大型的拟态辐射区。其中一个基因 ,已经在蝴蝶的颜色图案进化中被反复关联。在这里,我们在多个 物种中进行了 CRISPR 敲除实验,结果表明 是鳞片细胞身份的主要决定因素。染色质可及性分析和基因渐渗扫描确定了与离散表型开关相关的 -调控区域。在黑后翅基因型中,对这些区域进行 CRISPR 干扰重现了一个黄色条纹,揭示了它们空间上的局限性活性。在 谱系中,候选的黄色条纹表型变体的 CRE 被一个转座元件中断,这表明 -调控结构变异是这些拟态适应的基础。我们的工作表明, 功能上控制着鳞片的颜色命运,其 -调控区域以模块化和图案特异性的方式控制着表型开关。