Department of Structural Dynamics, MPI for Biophysical Chemistry, Göttingen, Germany.
Cellular Biochemistry, MPI for Biophysical Chemistry, Göttingen, Germany.
Nature. 2021 Aug;596(7871):296-300. doi: 10.1038/s41586-021-03789-5. Epub 2021 Aug 4.
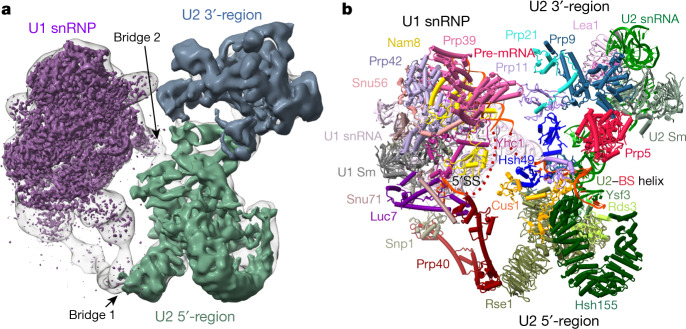
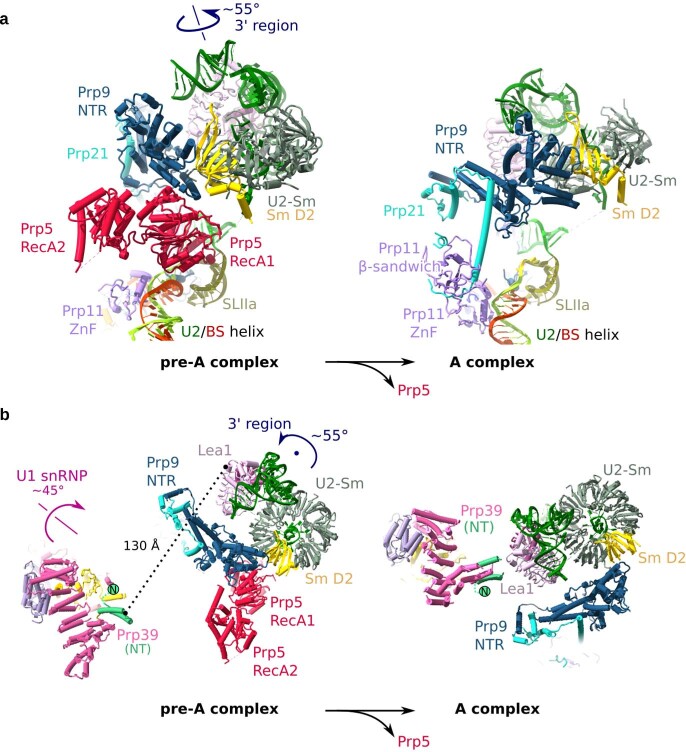
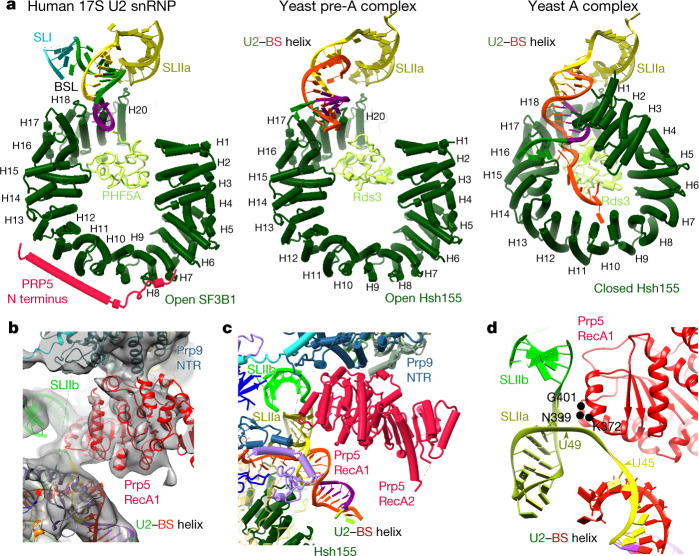
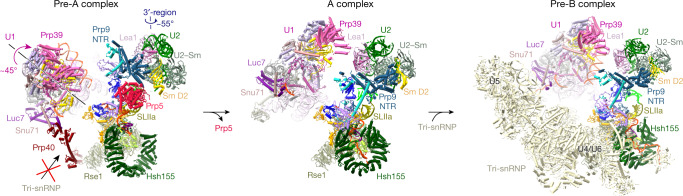
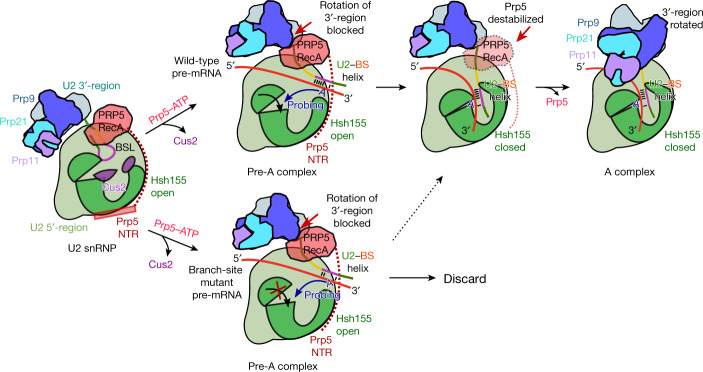
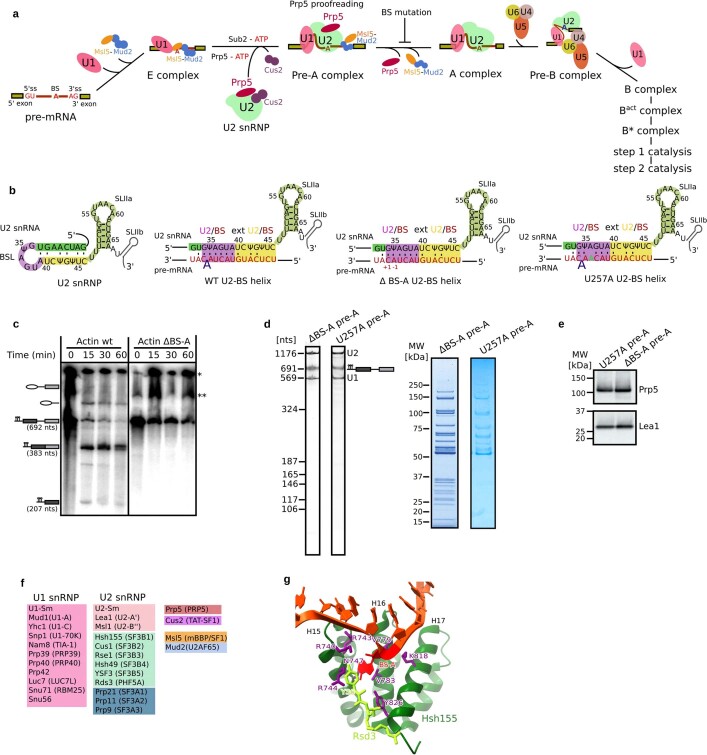
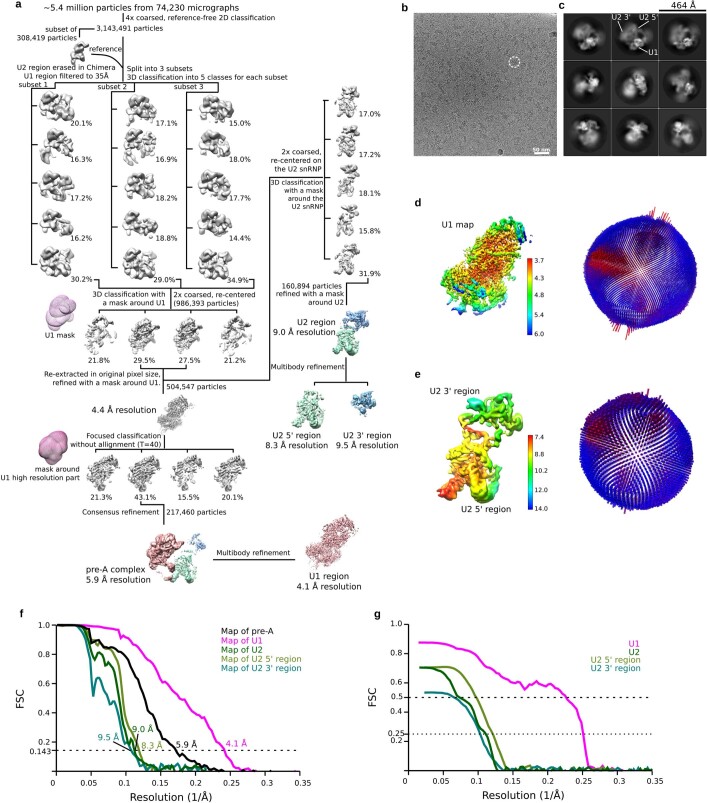
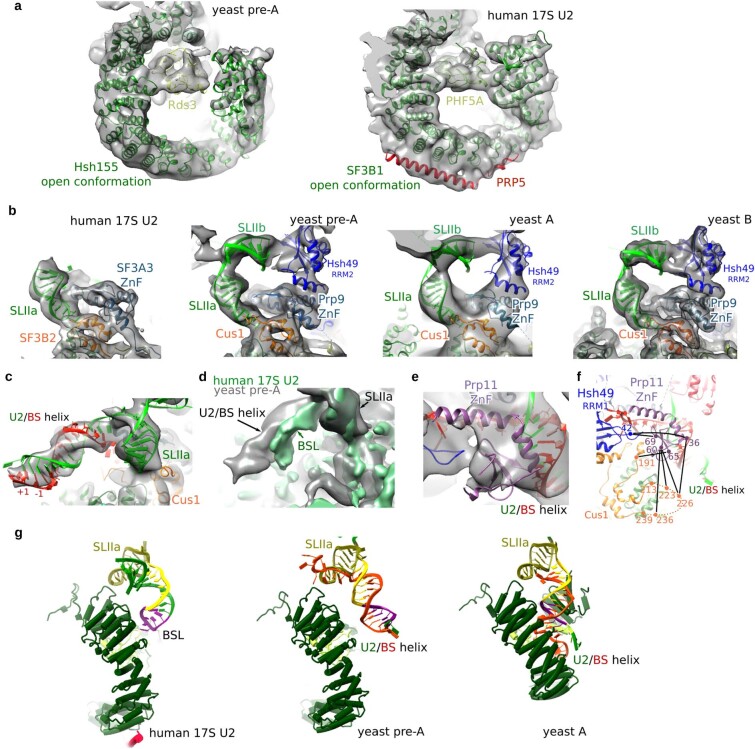
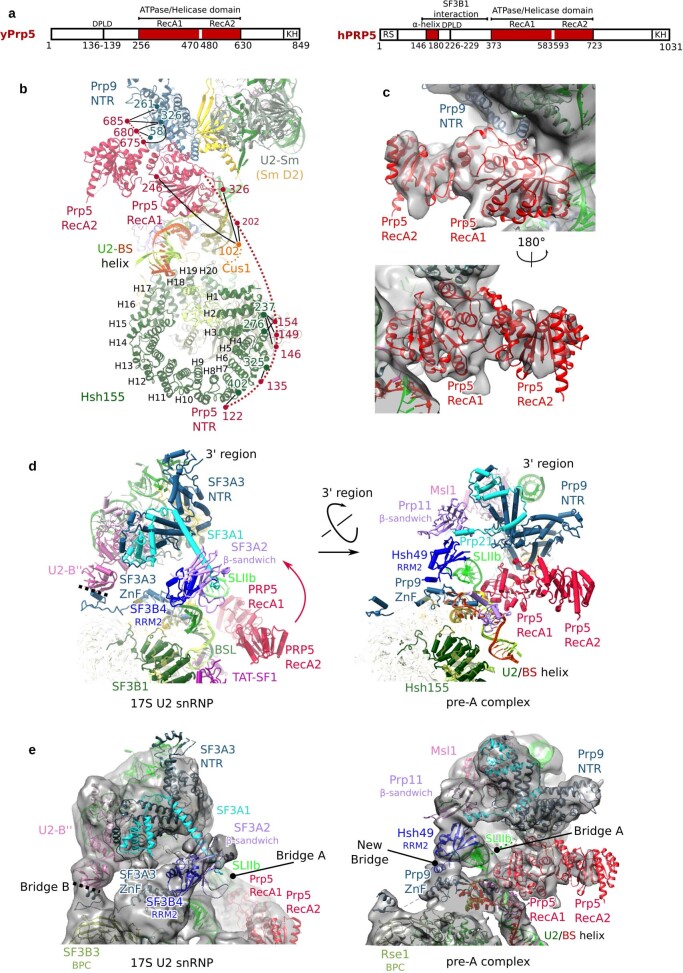
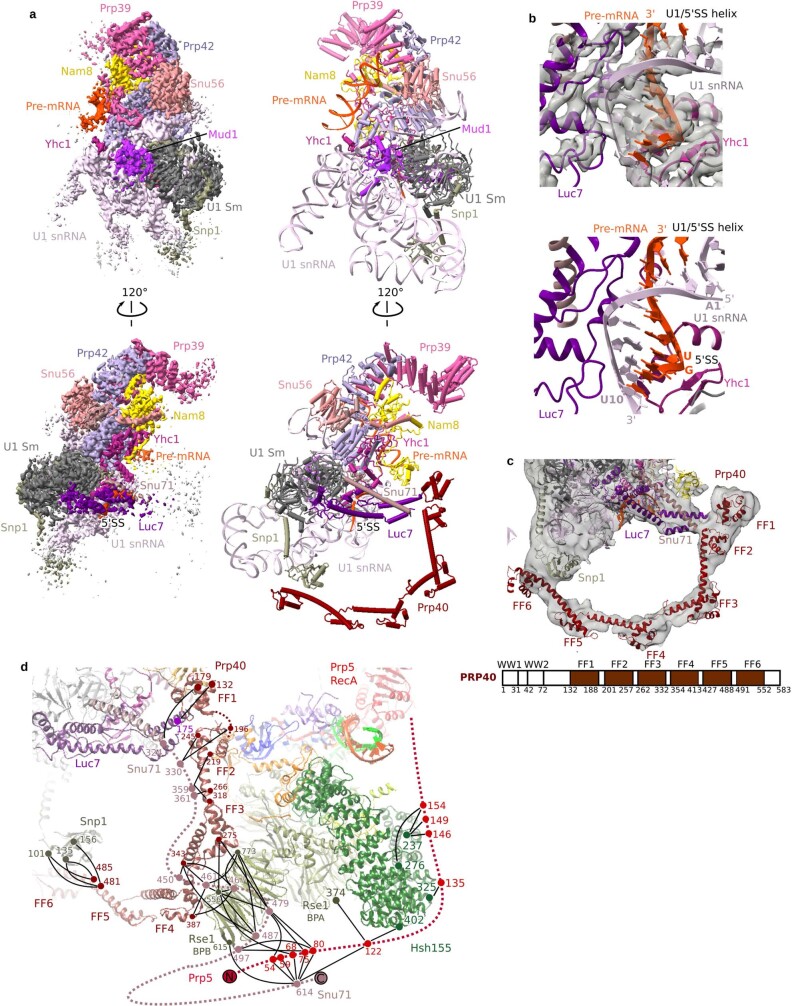
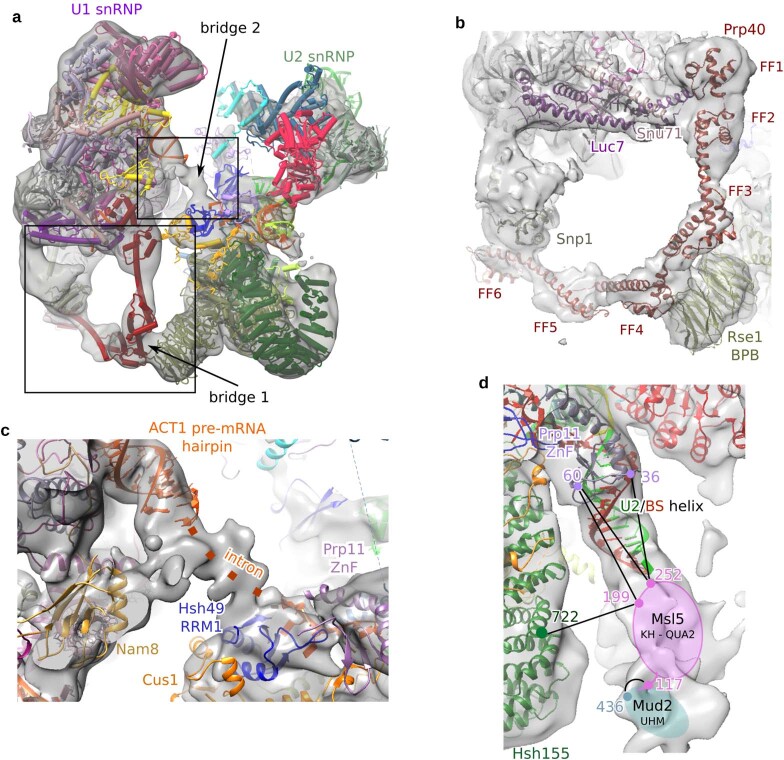
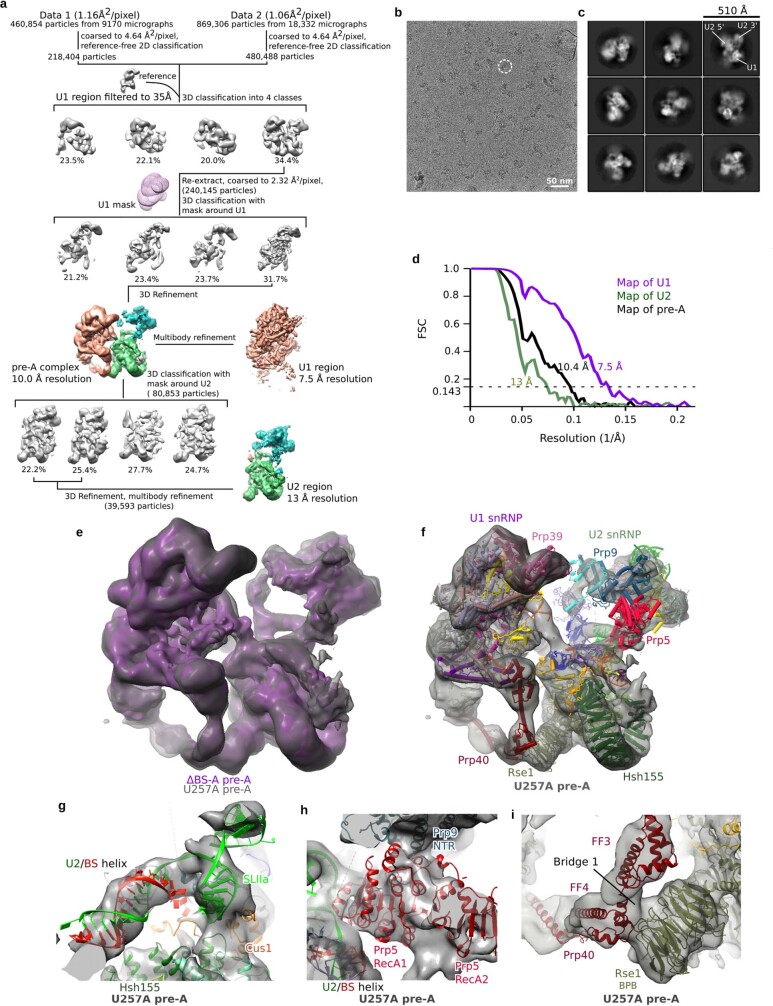
During the splicing of introns from precursor messenger RNAs (pre-mRNAs), the U2 small nuclear ribonucleoprotein (snRNP) must undergo stable integration into the spliceosomal A complex-a poorly understood, multistep process that is facilitated by the DEAD-box helicase Prp5 (refs. ). During this process, the U2 small nuclear RNA (snRNA) forms an RNA duplex with the pre-mRNA branch site (the U2-BS helix), which is proofread by Prp5 at this stage through an unclear mechanism. Here, by deleting the branch-site adenosine (BS-A) or mutating the branch-site sequence of an actin pre-mRNA, we stall the assembly of spliceosomes in extracts from the yeast Saccharomyces cerevisiae directly before the A complex is formed. We then determine the three-dimensional structure of this newly identified assembly intermediate by cryo-electron microscopy. Our structure indicates that the U2-BS helix has formed in this pre-A complex, but is not yet clamped by the HEAT domain of the Hsh155 protein (Hsh155), which exhibits an open conformation. The structure further reveals a large-scale remodelling/repositioning of the U1 and U2 snRNPs during the formation of the A complex that is required to allow subsequent binding of the U4/U6.U5 tri-snRNP, but that this repositioning is blocked in the pre-A complex by the presence of Prp5. Our data suggest that binding of Hsh155 to the bulged BS-A of the U2-BS helix triggers closure of Hsh155, which in turn destabilizes Prp5 binding. Thus, Prp5 proofreads the branch site indirectly, hindering spliceosome assembly if branch-site mutations prevent the remodelling of Hsh155. Our data provide structural insights into how a spliceosomal helicase enhances the fidelity of pre-mRNA splicing.
在将前体信使 RNA(pre-mRNA)中的内含子拼接过程中,U2 小核核糖核蛋白(snRNP)必须稳定地整合到剪接体 A 复合物中,这是一个理解甚少的多步骤过程,由 DEAD -box 解旋酶 Prp5 促进(参考文献)。在此过程中,U2 小核 RNA(snRNA)与前体 mRNA 分支位点(U2-BS 螺旋)形成 RNA 双链,Prp5 在这一阶段通过一个不明机制对其进行校对。在这里,通过删除分支位点腺苷(BS-A)或突变肌动蛋白 pre-mRNA 的分支位点序列,我们在酿酒酵母提取物中直接在 A 复合物形成之前阻止剪接体的组装。然后,我们通过冷冻电子显微镜确定了这个新鉴定的组装中间体的三维结构。我们的结构表明,U2-BS 螺旋已经在前 A 复合物中形成,但尚未被 Hsh155 蛋白(Hsh155)的 HEAT 结构域夹住,Hsh155 呈现开放构象。该结构进一步揭示了在 A 复合物形成过程中 U1 和 U2 snRNP 的大规模重排/重定位,这是允许随后结合 U4/U6.U5 三 snRNP 所必需的,但这种重定位在 Prp5 存在的情况下被前 A 复合物阻止。我们的数据表明,Hsh155 与 U2-BS 螺旋的膨出 BS-A 结合触发 Hsh155 的闭合,这反过来又使 Prp5 结合不稳定。因此,Prp5 间接地对分支位点进行校对,如果分支位点突变阻止 Hsh155 的重排,则阻碍剪接体组装。我们的数据为了解剪接体解旋酶如何提高 pre-mRNA 剪接的保真度提供了结构见解。