Department of Cell Biology, Harvard Medical School, Boston, Boston, United States.
Aligning Science Across Parkinson's (ASAP) Collaborative Research Network, Chevy Chase, United States.
Elife. 2021 Sep 29;10:e72328. doi: 10.7554/eLife.72328.
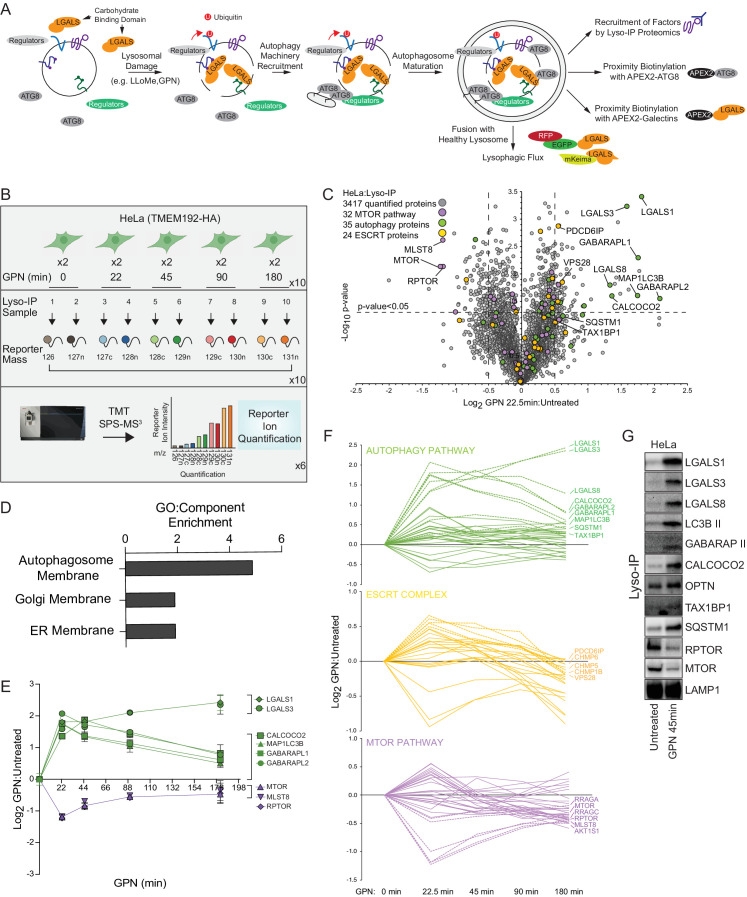
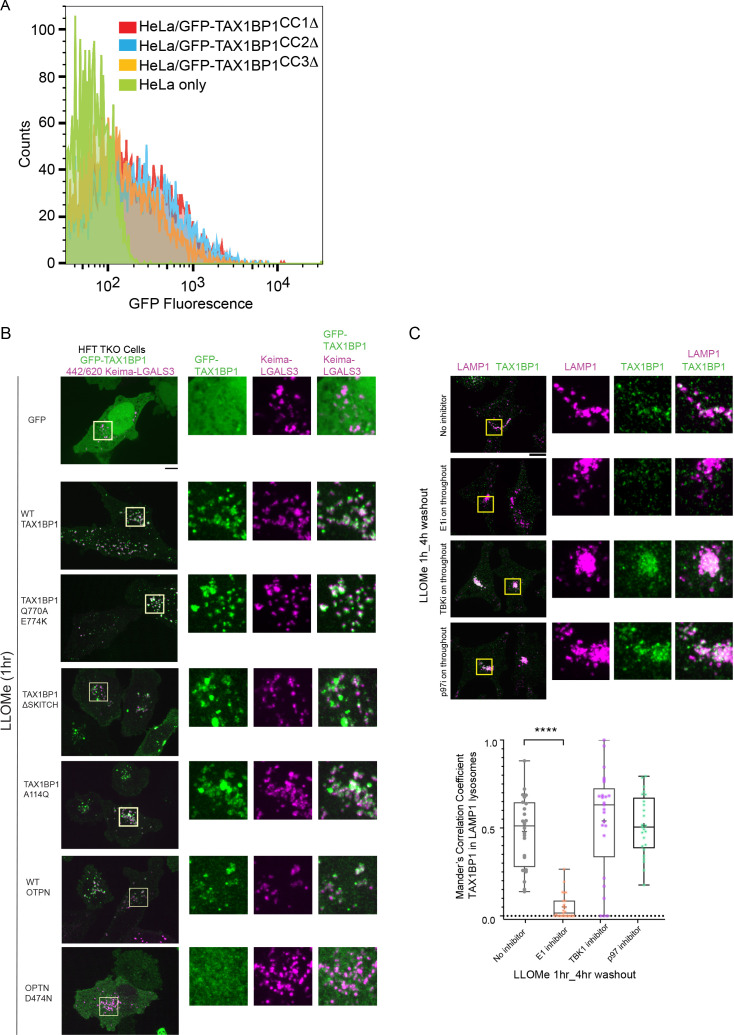
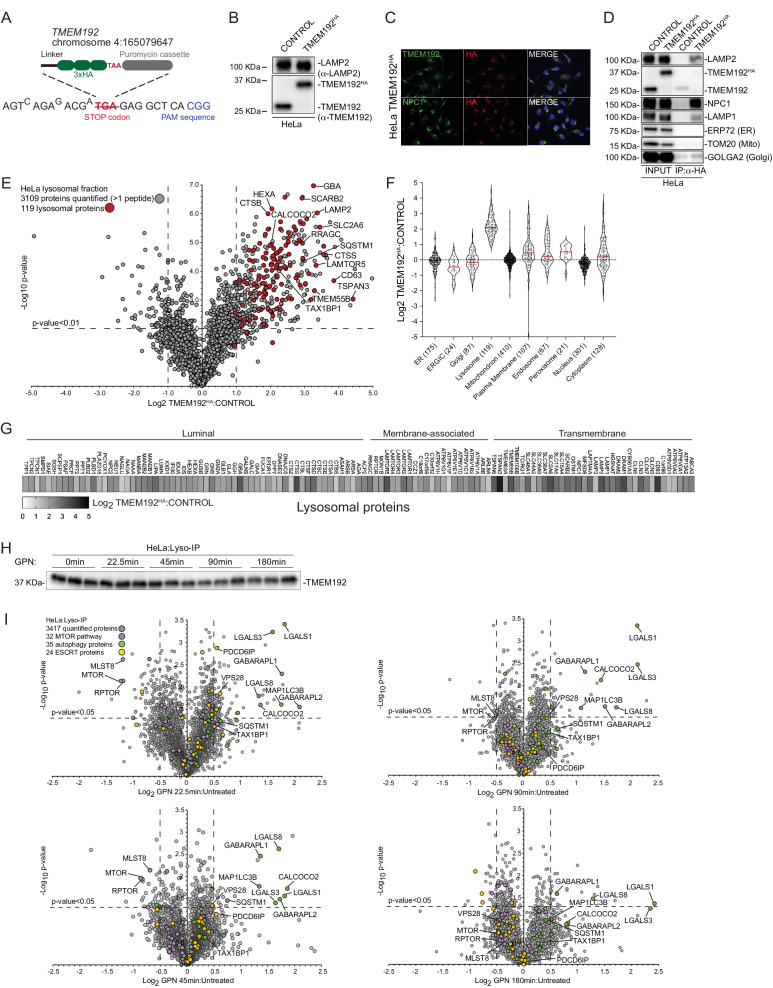
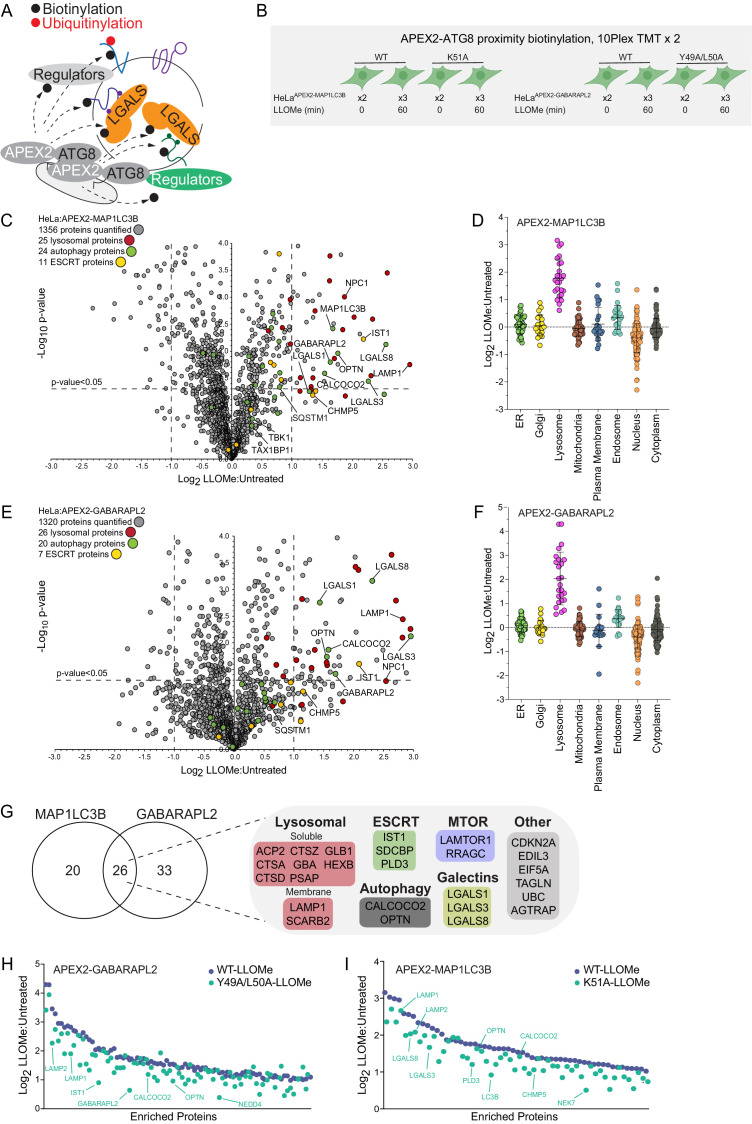
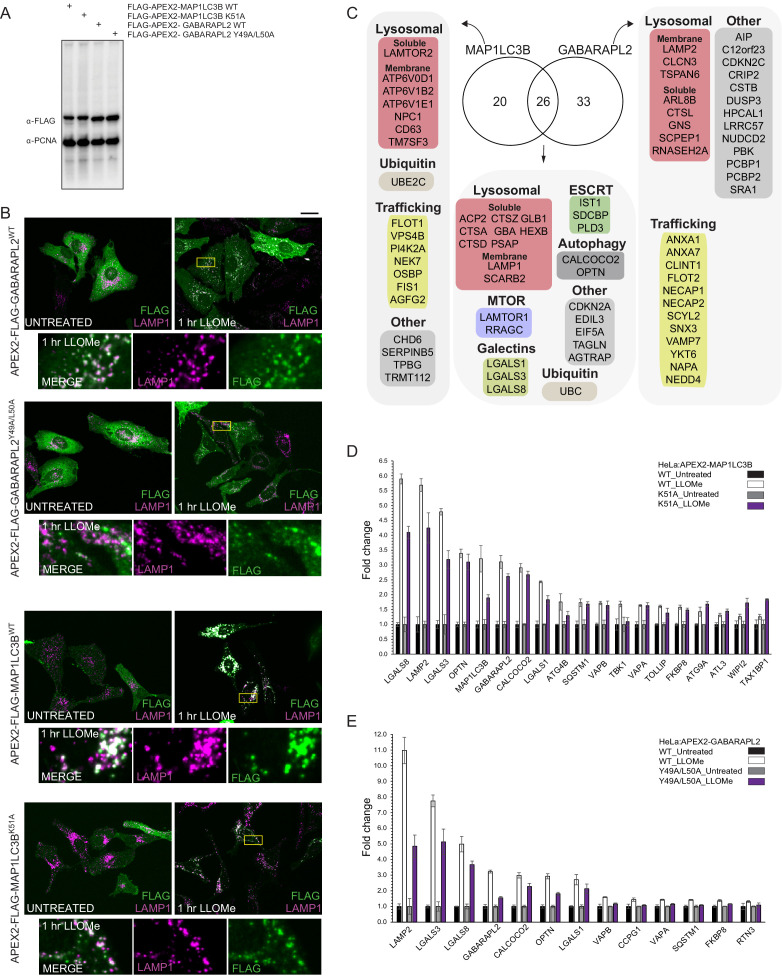
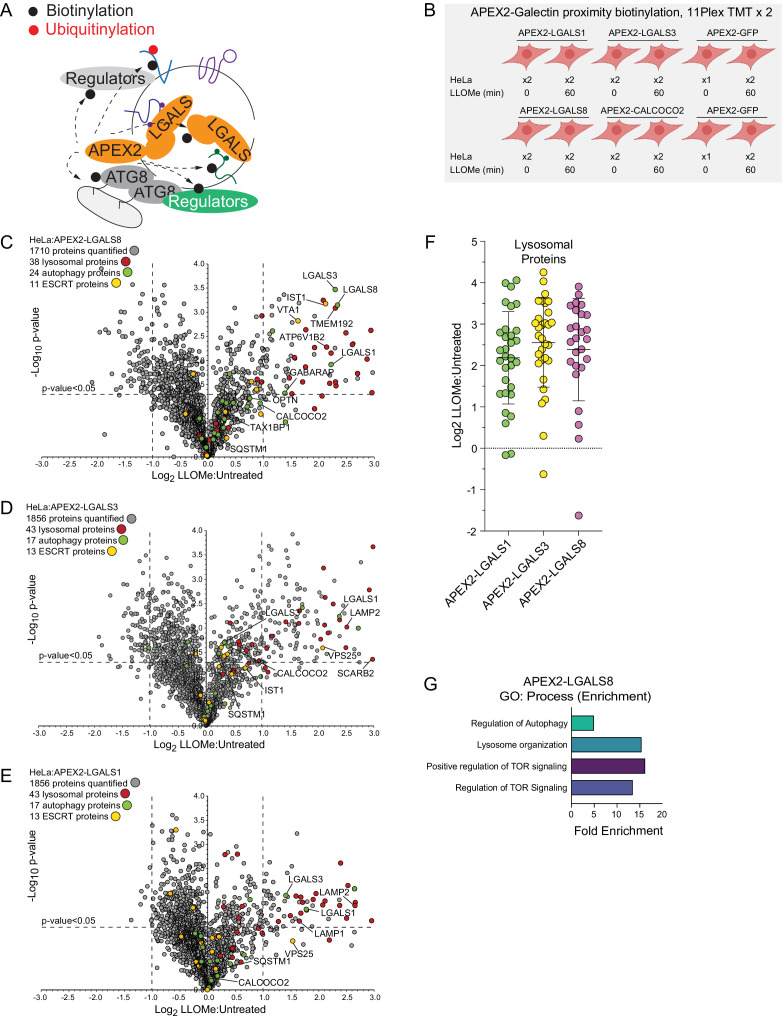
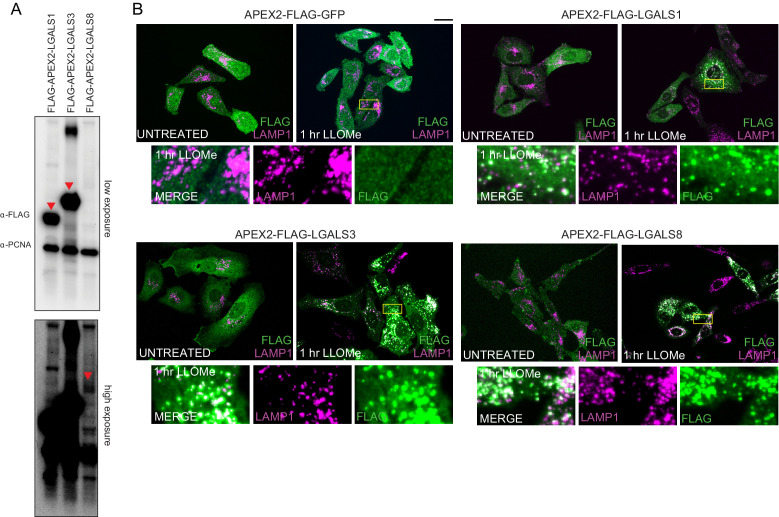
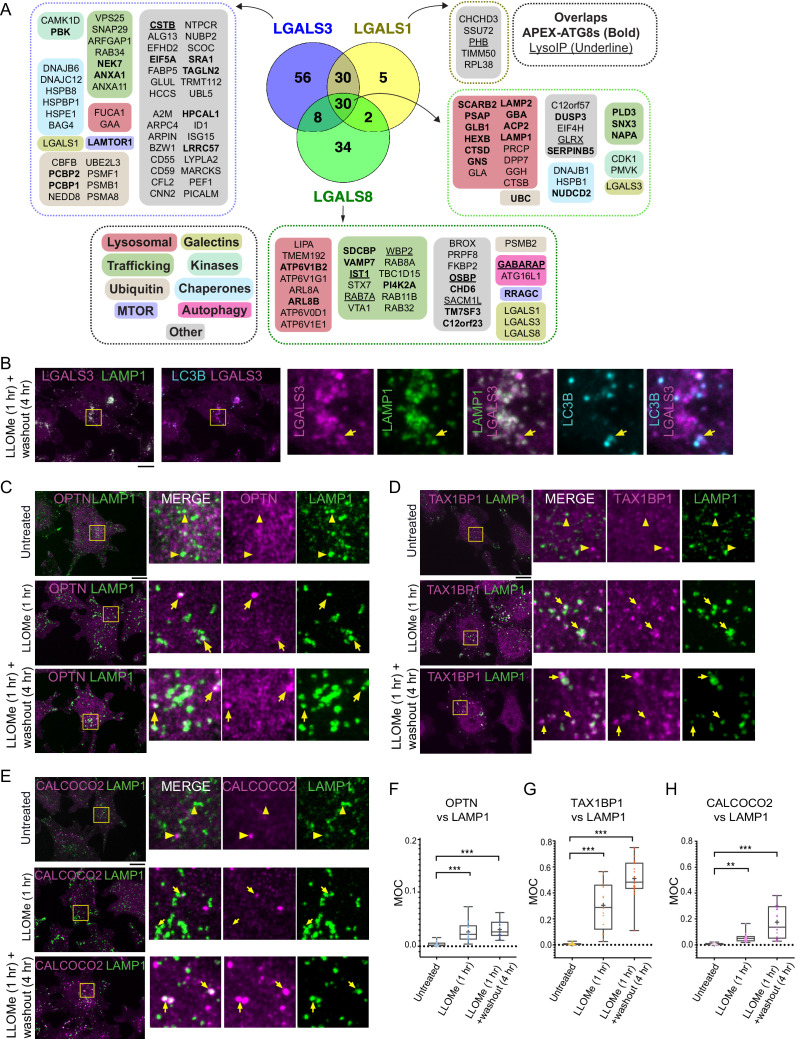
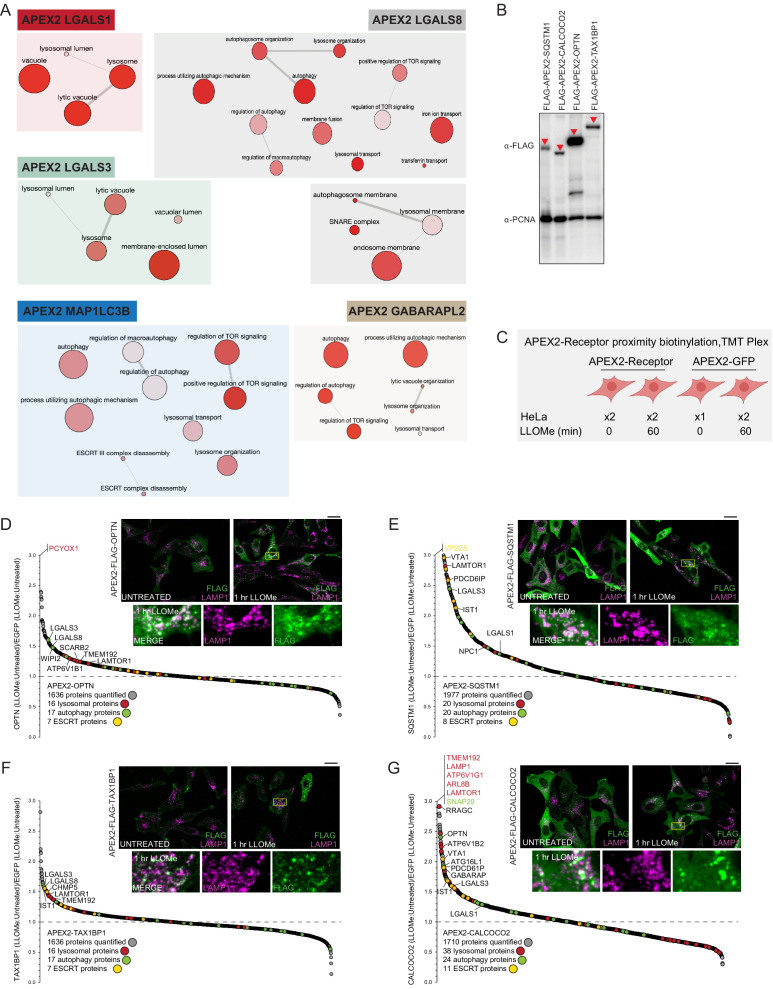
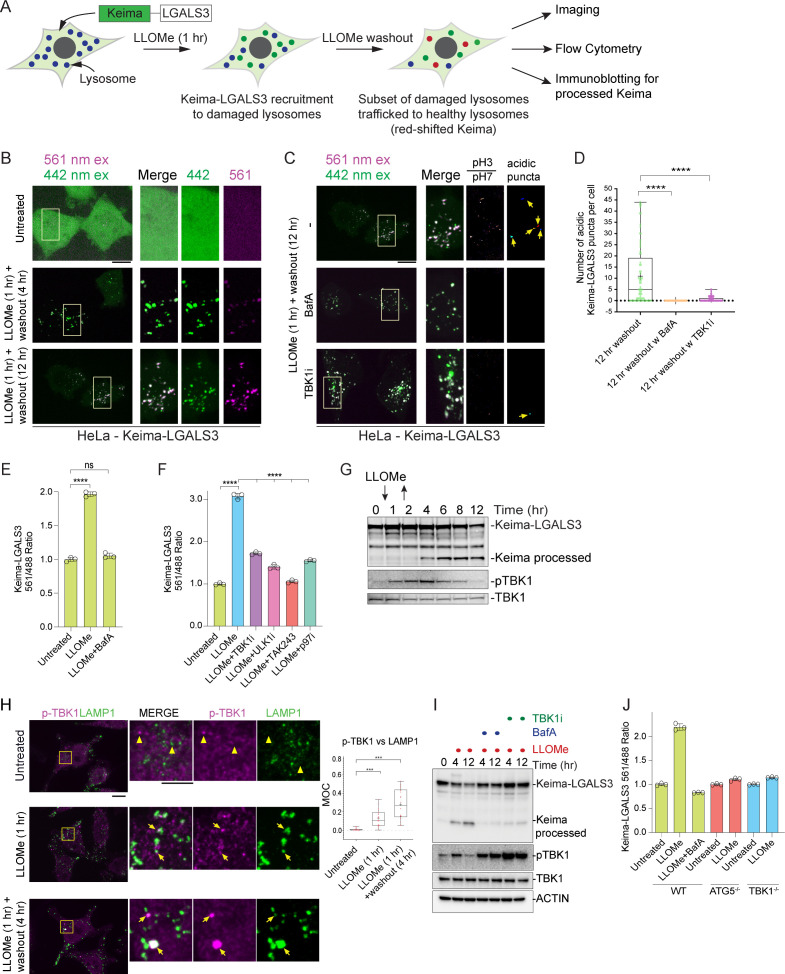
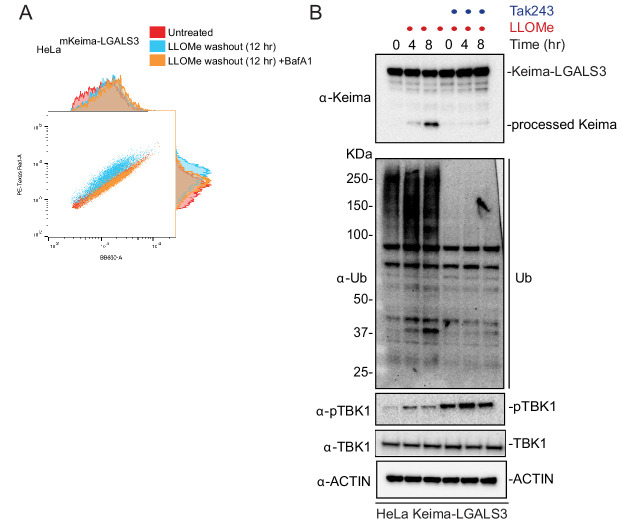
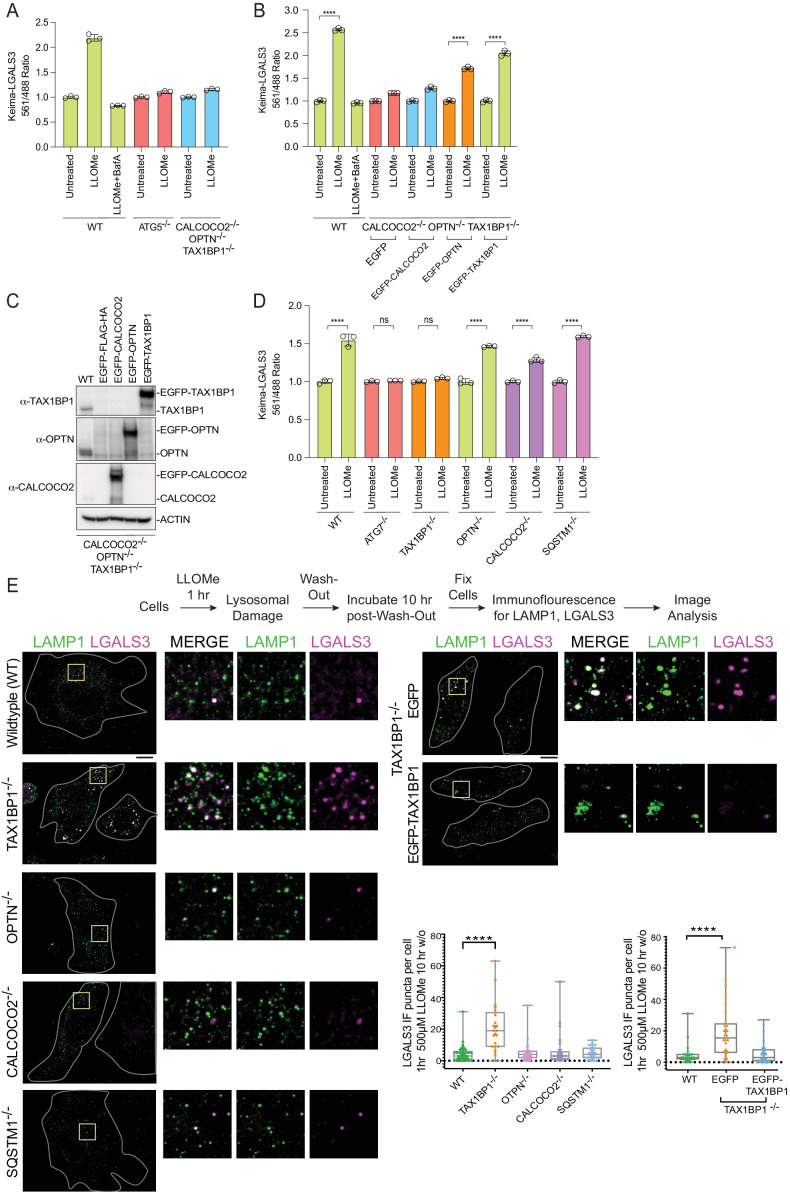
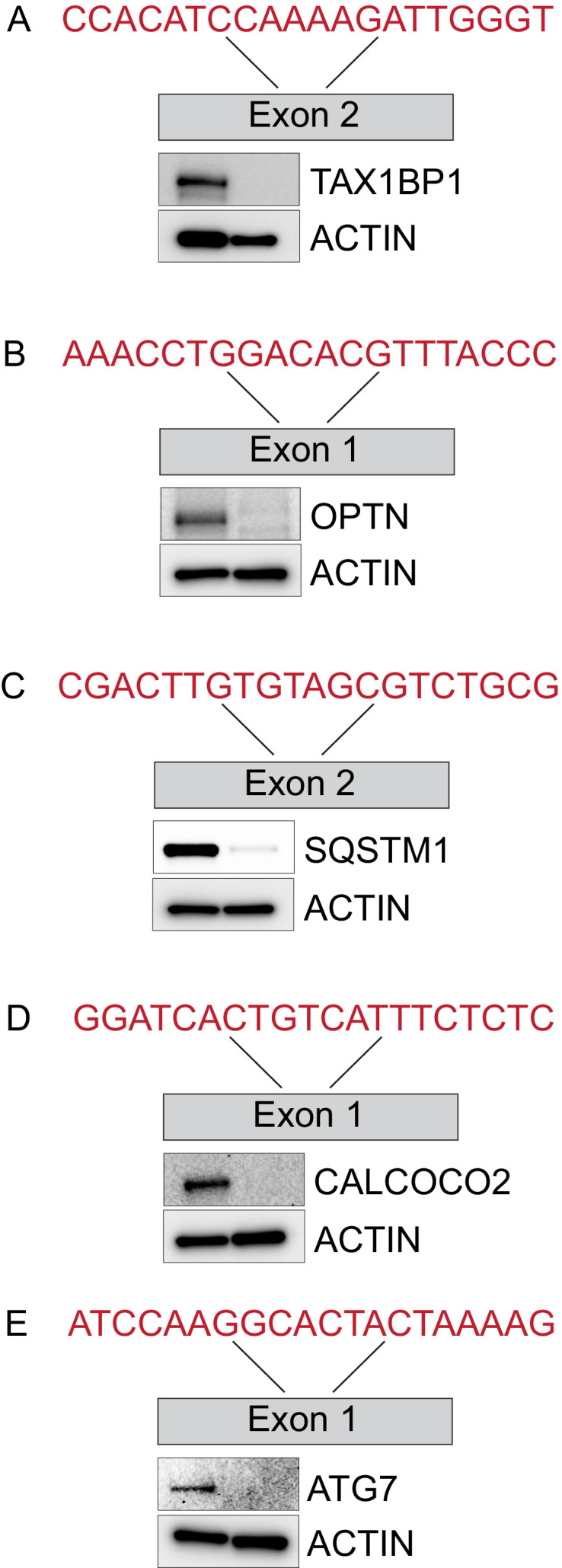
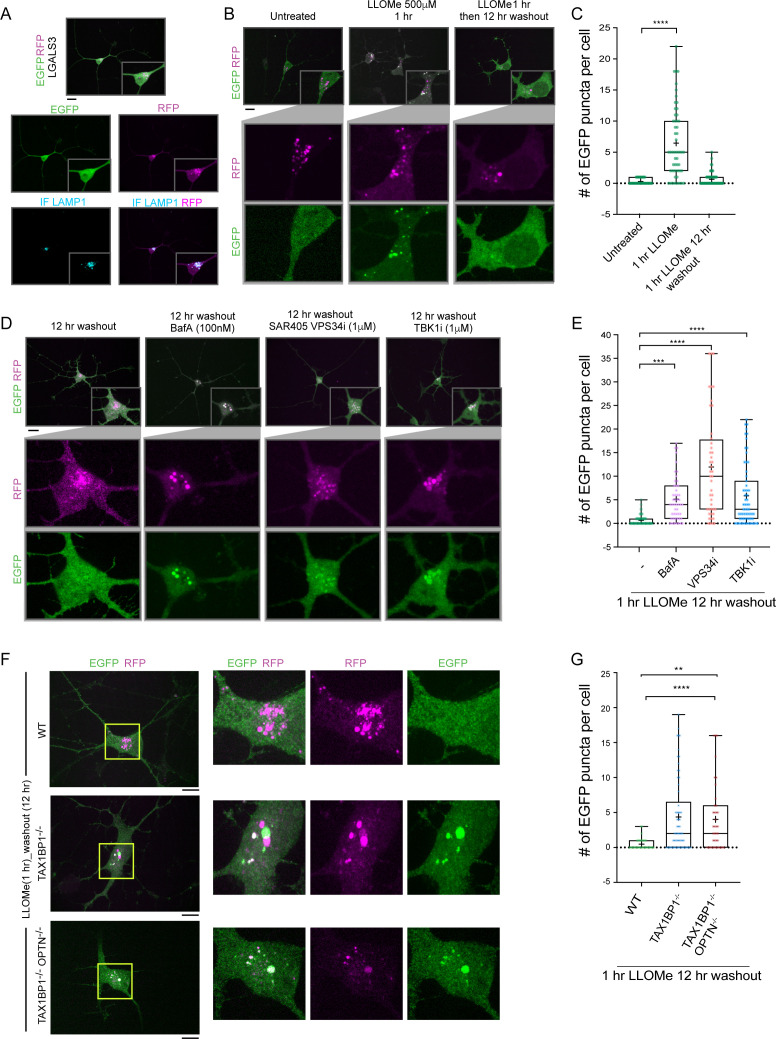
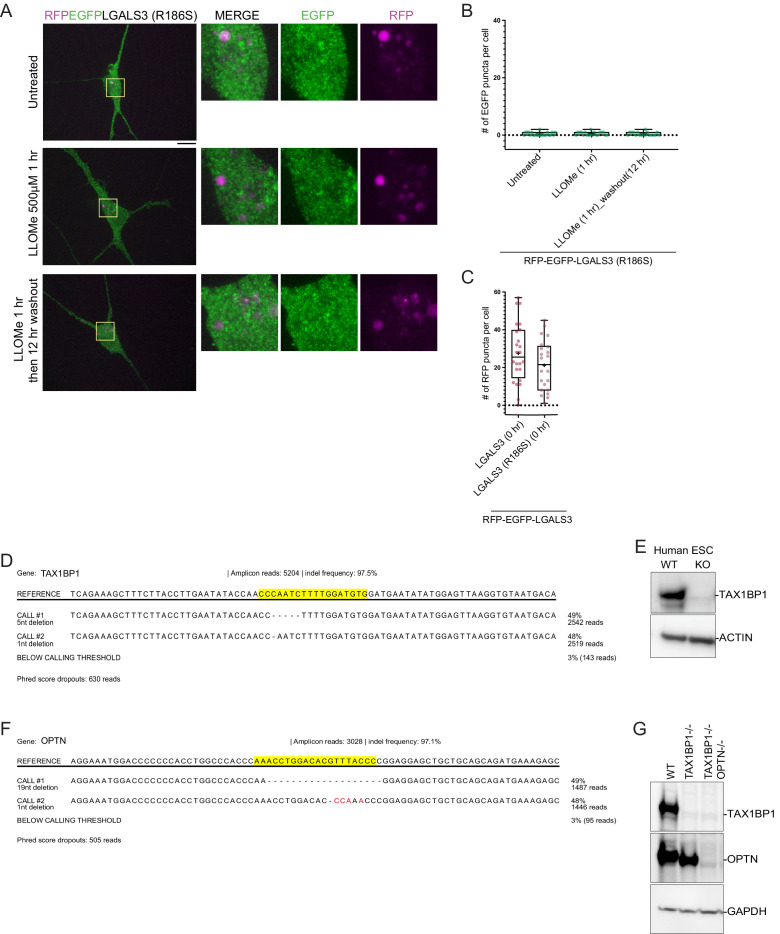
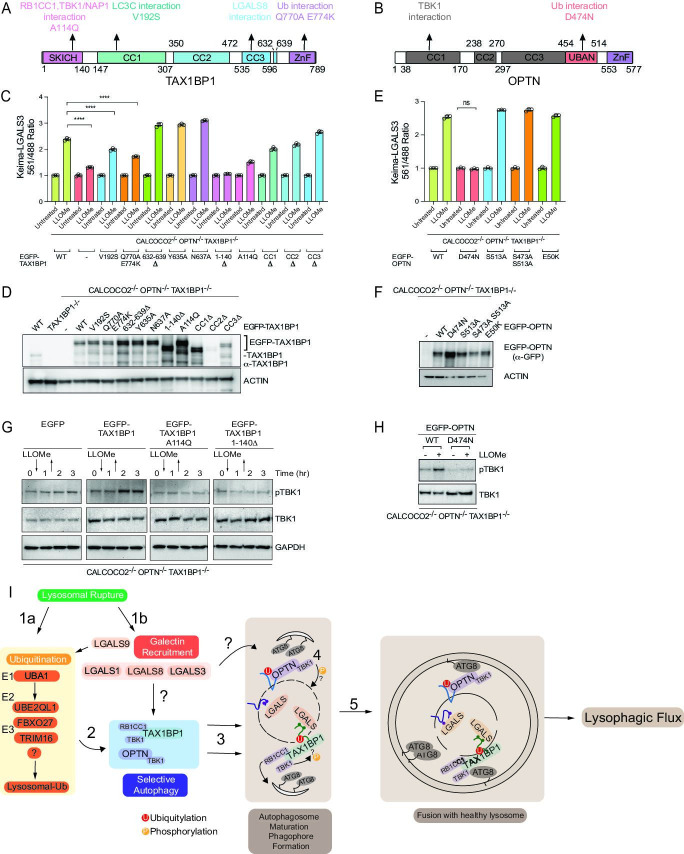
Removal of damaged organelles via the process of selective autophagy constitutes a major form of cellular quality control. Damaged organelles are recognized by a dedicated surveillance machinery, leading to the assembly of an autophagosome around the damaged organelle, prior to fusion with the degradative lysosomal compartment. Lysosomes themselves are also prone to damage and are degraded through the process of lysophagy. While early steps involve recognition of ruptured lysosomal membranes by glycan-binding galectins and ubiquitylation of transmembrane lysosomal proteins, many steps in the process, and their interrelationships, remain poorly understood, including the role and identity of cargo receptors required for completion of lysophagy. Here, we employ quantitative organelle capture and proximity biotinylation proteomics of autophagy adaptors, cargo receptors, and galectins in response to acute lysosomal damage, thereby revealing the landscape of lysosome-associated proteome remodeling during lysophagy. Among the proteins dynamically recruited to damaged lysosomes were ubiquitin-binding autophagic cargo receptors. Using newly developed lysophagic flux reporters including Lyso-Keima, we demonstrate that TAX1BP1, together with its associated kinase TBK1, are both necessary and sufficient to promote lysophagic flux in both HeLa cells and induced neurons (iNeurons). While the related receptor Optineurin (OPTN) can drive damage-dependent lysophagy when overexpressed, cells lacking either OPTN or CALCOCO2 still maintain significant lysophagic flux in HeLa cells. Mechanistically, TAX1BP1-driven lysophagy requires its N-terminal SKICH domain, which binds both TBK1 and the autophagy regulatory factor RB1CC1, and requires upstream ubiquitylation events for efficient recruitment and lysophagic flux. These results identify TAX1BP1 as a central component in the lysophagy pathway and provide a proteomic resource for future studies of the lysophagy process.
通过选择性自噬过程清除受损细胞器是细胞质量控制的主要形式。受损细胞器被专门的监测机制识别,导致在与降解性溶酶体隔室融合之前,在受损细胞器周围组装自噬体。溶酶体本身也容易受到损伤,并通过溶酶体噬作用降解。虽然早期步骤涉及糖结合半乳糖凝集素识别破裂的溶酶体膜和跨膜溶酶体蛋白的泛素化,但该过程的许多步骤及其相互关系仍知之甚少,包括完成溶酶体噬作用所需的货物受体的作用和身份。在这里,我们通过定量细胞器捕获和自噬衔接子、货物受体和半乳糖凝集素的邻近生物素化蛋白质组学研究急性溶酶体损伤,从而揭示了溶酶体相关蛋白质组在溶酶体噬作用过程中的重塑景观。在动态募集到受损溶酶体的蛋白质中,有泛素结合的自噬货物受体。使用新开发的包括 Lyso-Keima 在内的溶酶体噬作用通量报告器,我们证明 TAX1BP1 与其相关激酶 TBK1 都是促进 HeLa 细胞和诱导神经元(iNeurons)中溶酶体噬作用通量所必需且充分的。虽然相关受体 Optineurin(OPTN)在过表达时可以驱动损伤依赖性的溶酶体噬作用,但缺乏 OPTN 或 CALCOCO2 的细胞在 HeLa 细胞中仍保持显著的溶酶体噬作用通量。从机制上讲,TAX1BP1 驱动的溶酶体噬作用需要其 N 端 SKICH 结构域,该结构域与 TBK1 和自噬调节因子 RB1CC1 结合,并需要上游泛素化事件来有效募集和溶酶体噬作用通量。这些结果确定 TAX1BP1 为溶酶体噬作用途径的核心组成部分,并为未来的溶酶体噬作用过程研究提供了蛋白质组资源。