Instituto de Biología Molecular y Celular de Plantas (CSIC-Universidad Politécnica de Valencia), 46022 Valencia, Spain.
Dipartimento di Bioscienze, Università degli Studi di Milano, 20133 Milano, Italy.
Plant Physiol. 2021 Nov 3;187(3):1534-1550. doi: 10.1093/plphys/kiab348.
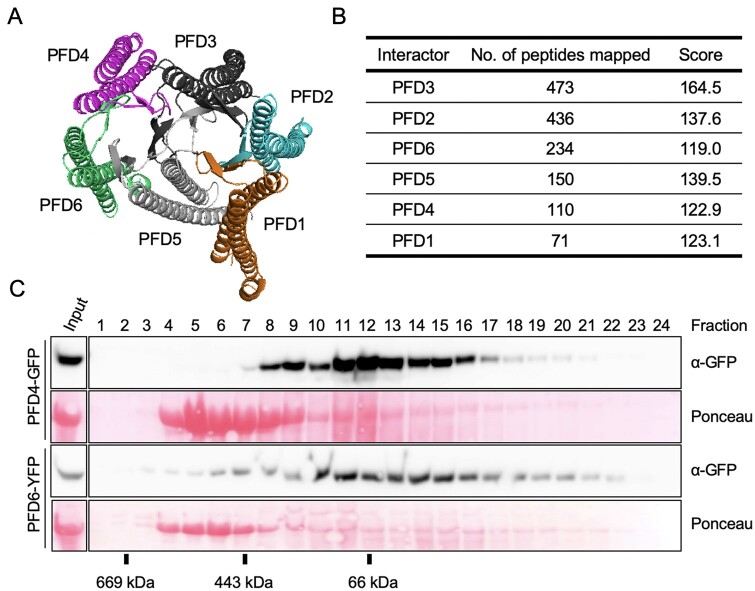
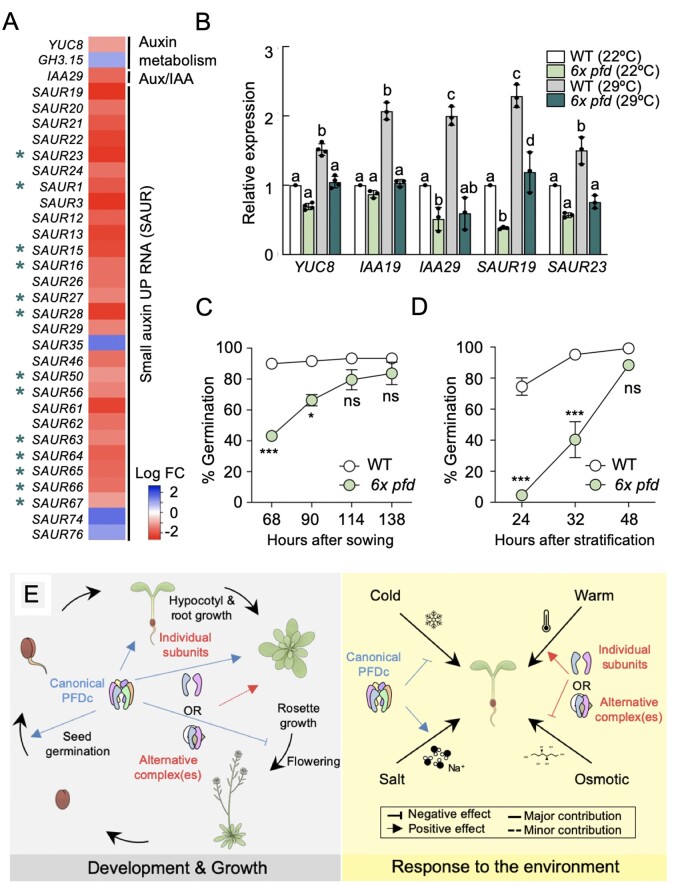
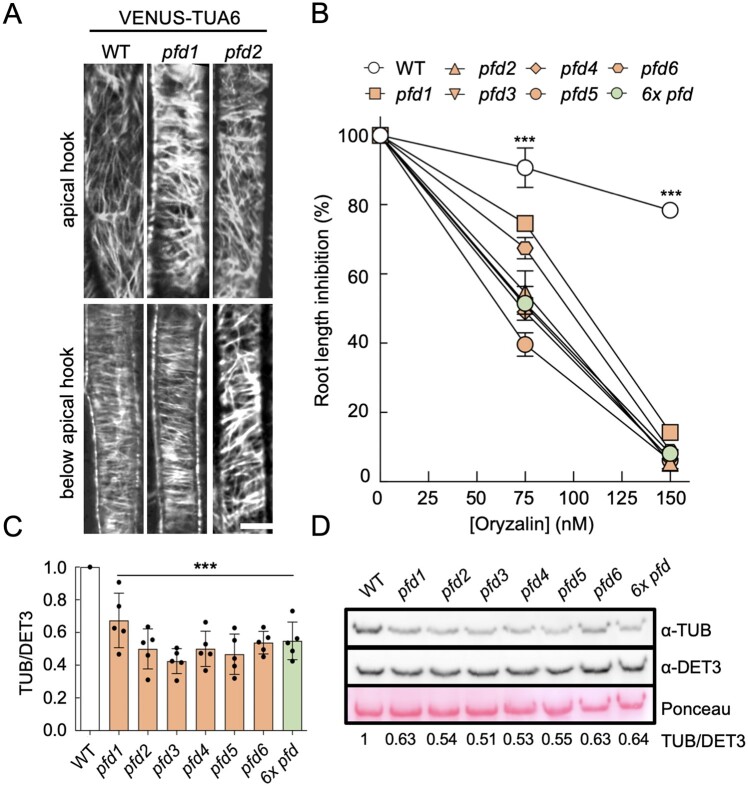
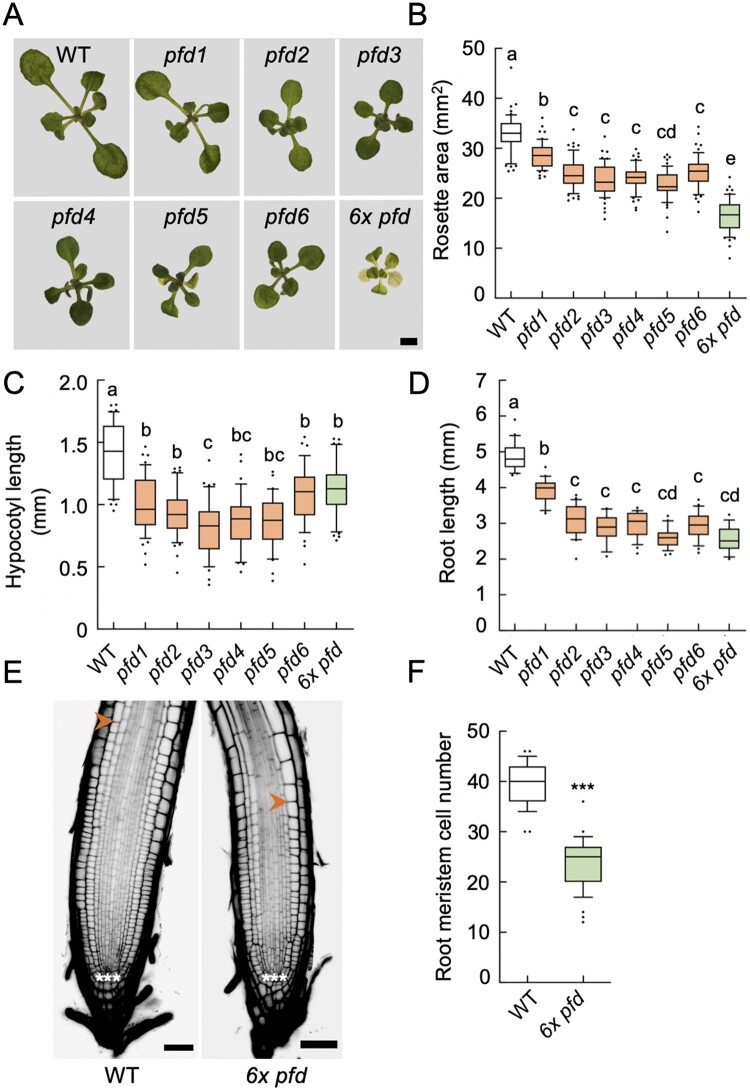
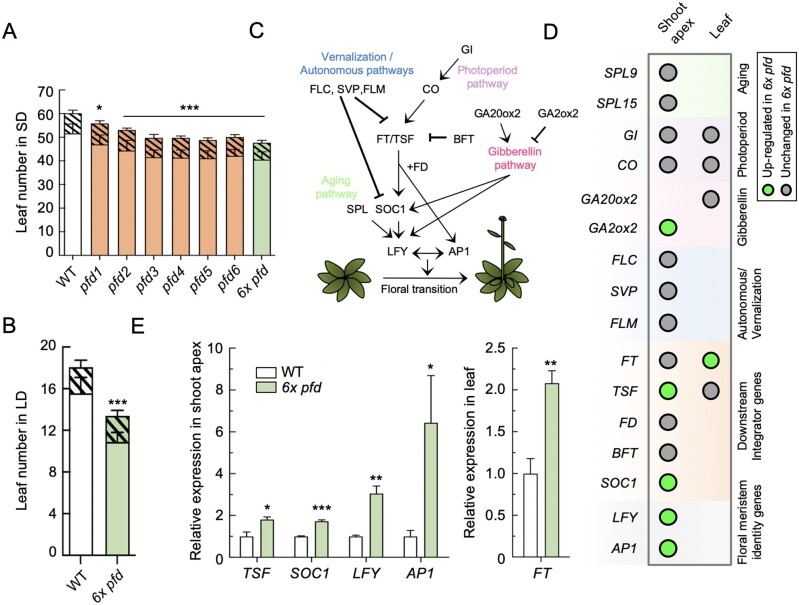
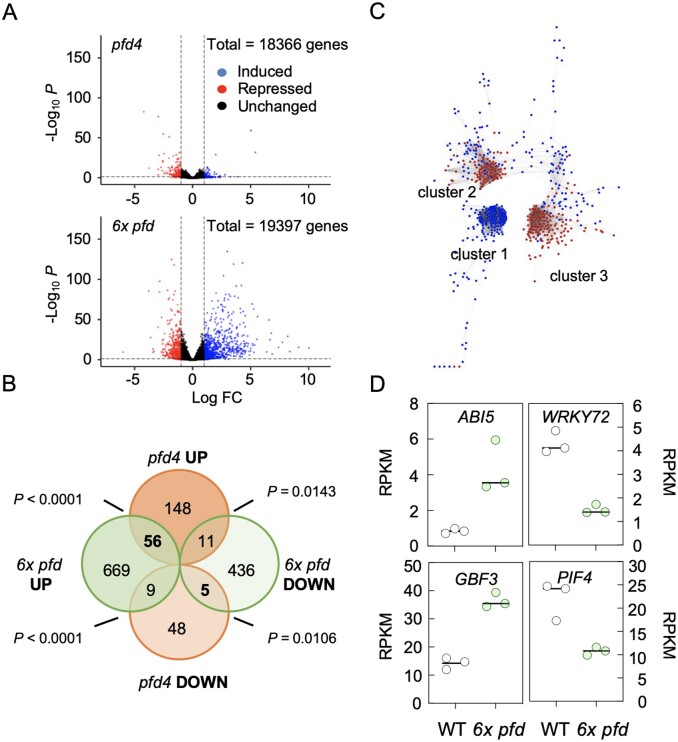
The prefoldin complex (PFDc) was identified in humans as a co-chaperone of the cytosolic chaperonin T-COMPLEX PROTEIN RING COMPLEX (TRiC)/CHAPERONIN CONTAINING TCP-1 (CCT). PFDc is conserved in eukaryotes and is composed of subunits PFD1-6, and PFDc-TRiC/CCT folds actin and tubulins. PFDs also participate in a wide range of cellular processes, both in the cytoplasm and in the nucleus, and their malfunction causes developmental alterations and disease in animals and altered growth and environmental responses in yeast and plants. Genetic analyses in yeast indicate that not all of their functions require the canonical complex. The lack of systematic genetic analyses in plants and animals, however, makes it difficult to discern whether PFDs participate in a process as the canonical complex or in alternative configurations, which is necessary to understand their mode of action. To tackle this question, and on the premise that the canonical complex cannot be formed if one subunit is missing, we generated an Arabidopsis (Arabidopsis thaliana) mutant deficient in the six PFDs and compared various growth and environmental responses with those of the individual mutants. In this way, we demonstrate that the PFDc is required for seed germination, to delay flowering, or to respond to high salt stress or low temperature, whereas at least two PFDs redundantly attenuate the response to osmotic stress. A coexpression analysis of differentially expressed genes in the sextuple mutant identified several transcription factors, including ABA INSENSITIVE 5 (ABI5) and PHYTOCHROME-INTERACTING FACTOR 4, acting downstream of PFDs. Furthermore, the transcriptomic analysis allowed assigning additional roles for PFDs, for instance, in response to higher temperature.
原核起始因子 3 亚基 F(IF3F)是真核生物 IF3 的同源物,其在原核生物和真核生物中都发挥着重要的功能。IF3F 与 30S 核糖体亚基结合,并与 IF1 和 IF2 相互作用,参与了翻译的起始过程。IF3F 在原核生物中与 IF1 和 IF2 形成 IF3,该复合物与 30S 核糖体结合,促进了 mRNA 与核糖体的结合。IF3F 在真核生物中也与 IF1 和 IF2 形成 IF3,该复合物参与了 40S 前核糖体的形成和起始复合物的组装。IF3F 在细胞内的表达和活性受到多种因素的调节,包括翻译起始因子的磷酸化、mRNA 的二级结构和翻译起始因子的相互作用等。IF3F 的突变或表达异常与多种疾病的发生和发展有关,如癌症、神经退行性疾病和病毒感染等。