Institute for Developmental Genetics and Cluster of Excellence on Plant Sciences, Heinrich Heine University, Düsseldorf, Germany.
Elife. 2021 Oct 13;10:e70934. doi: 10.7554/eLife.70934.
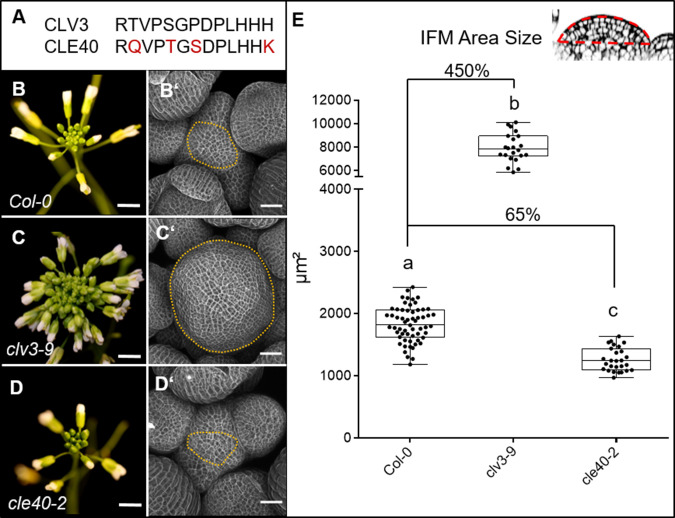
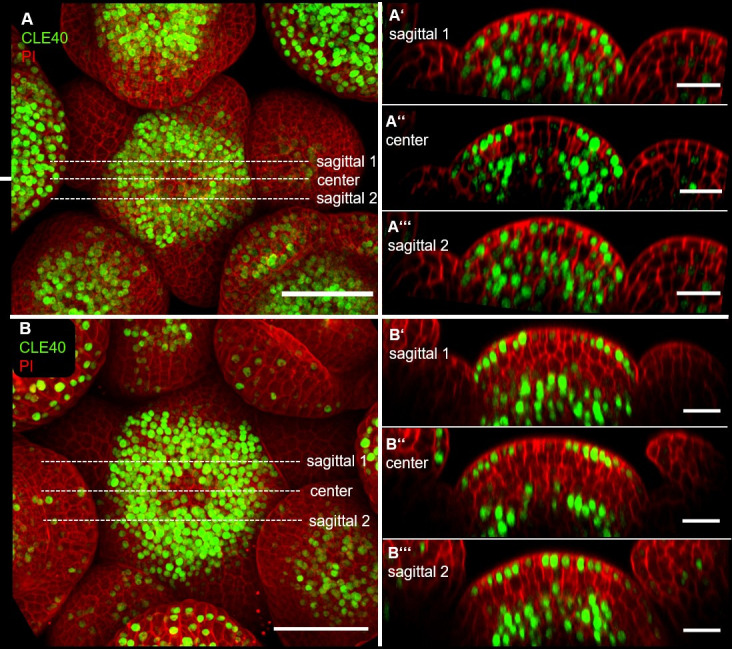
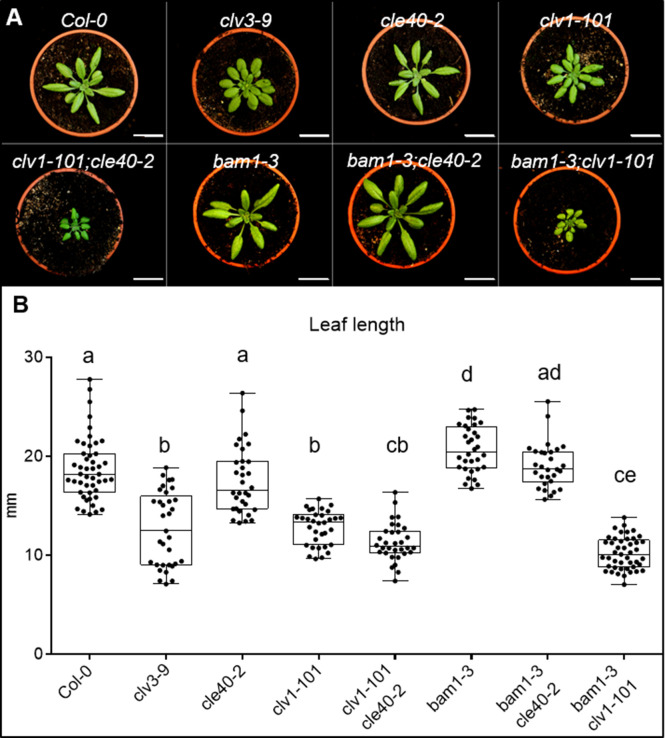
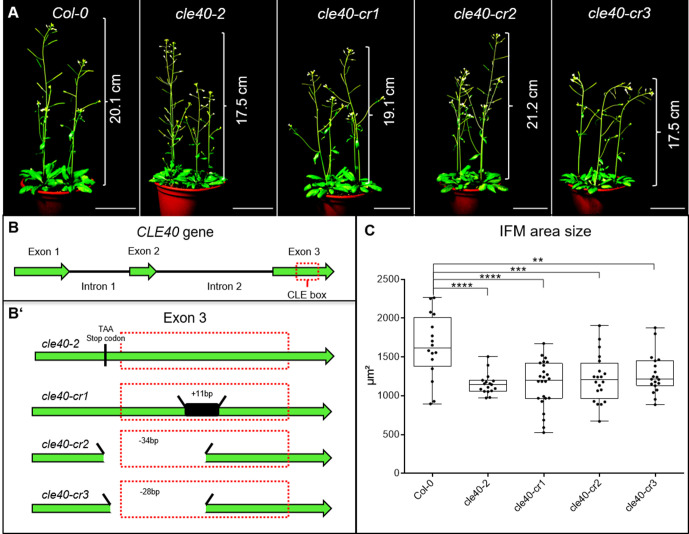
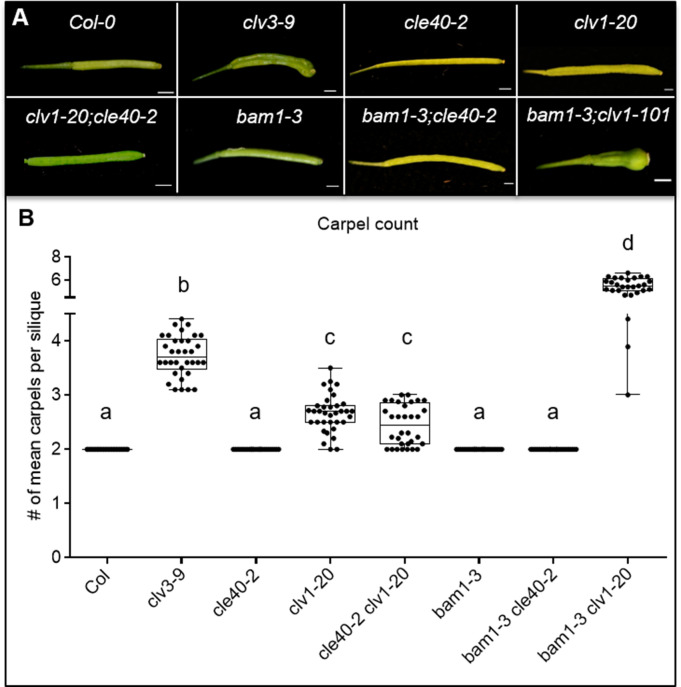
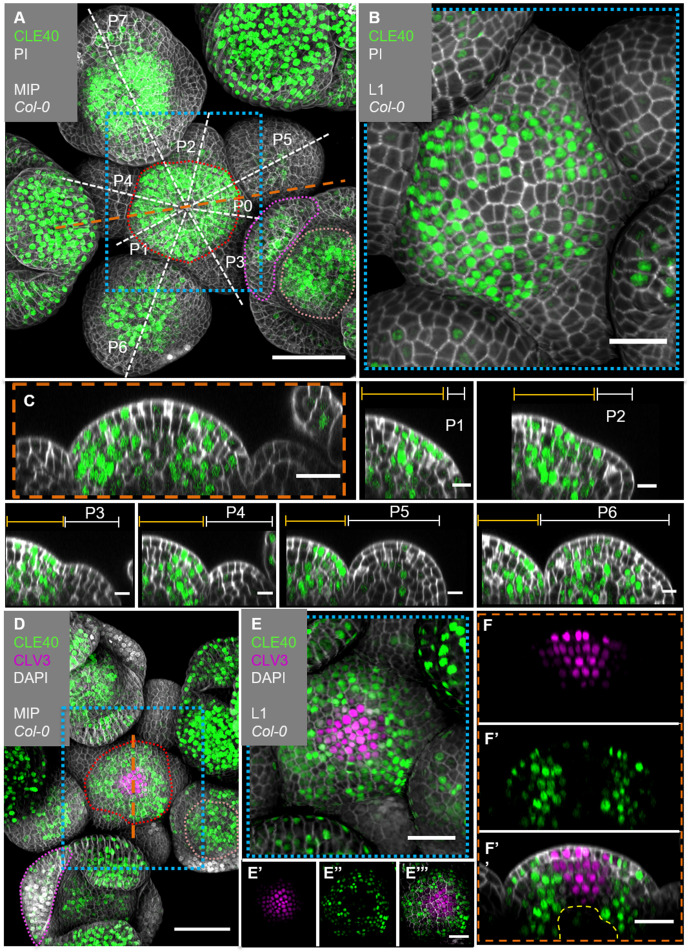
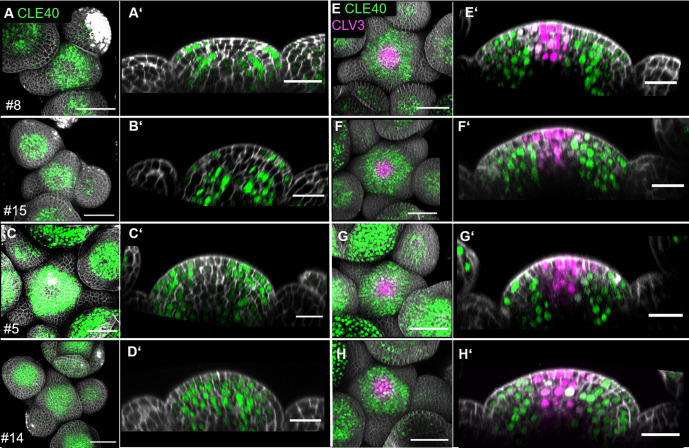
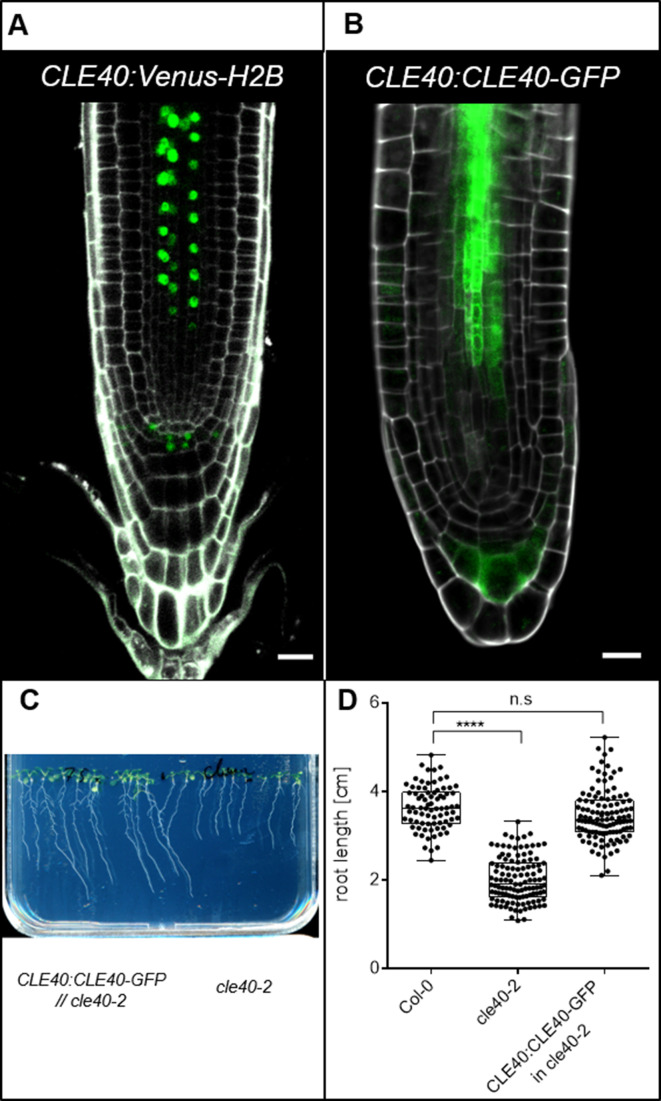
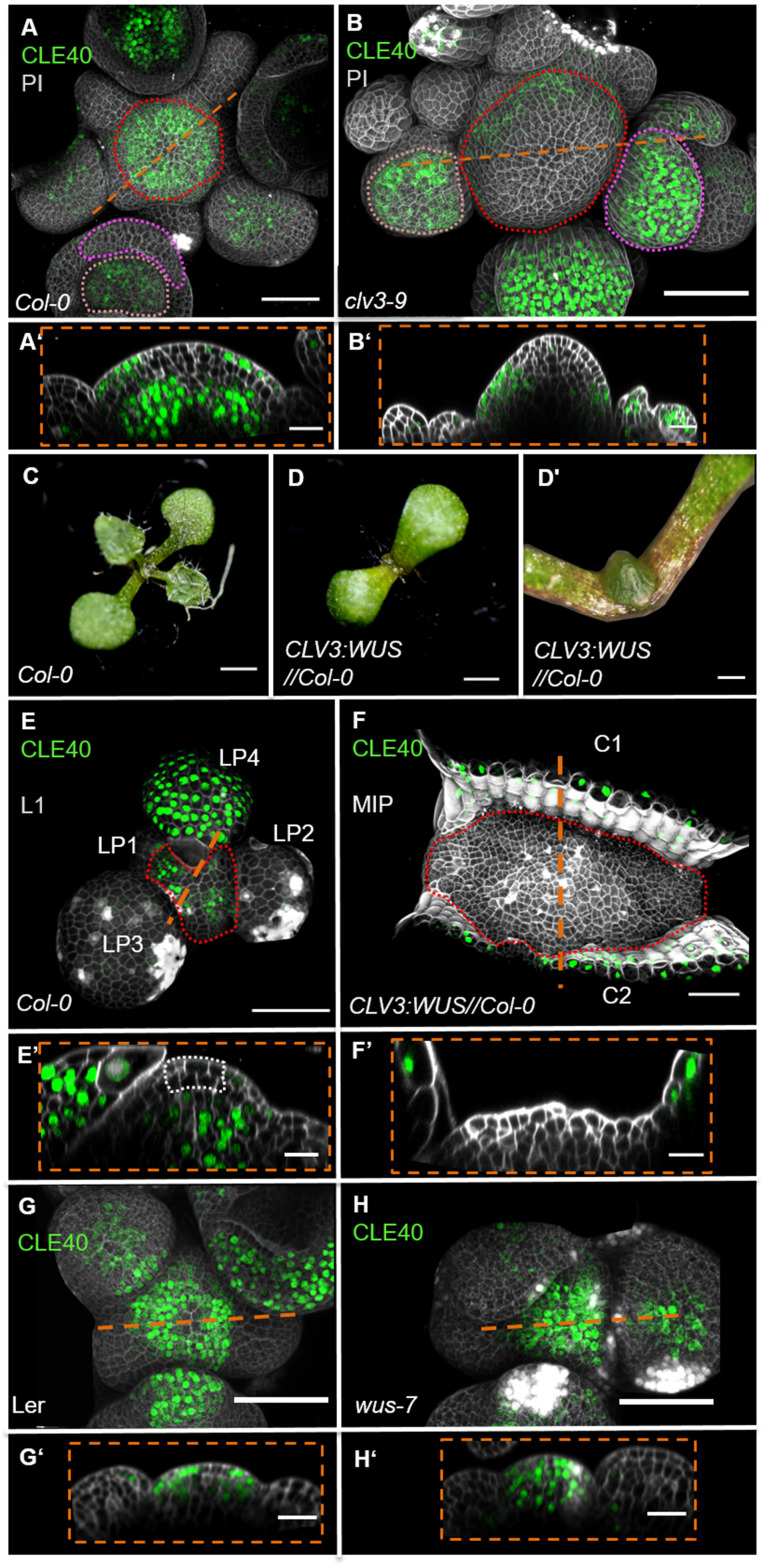
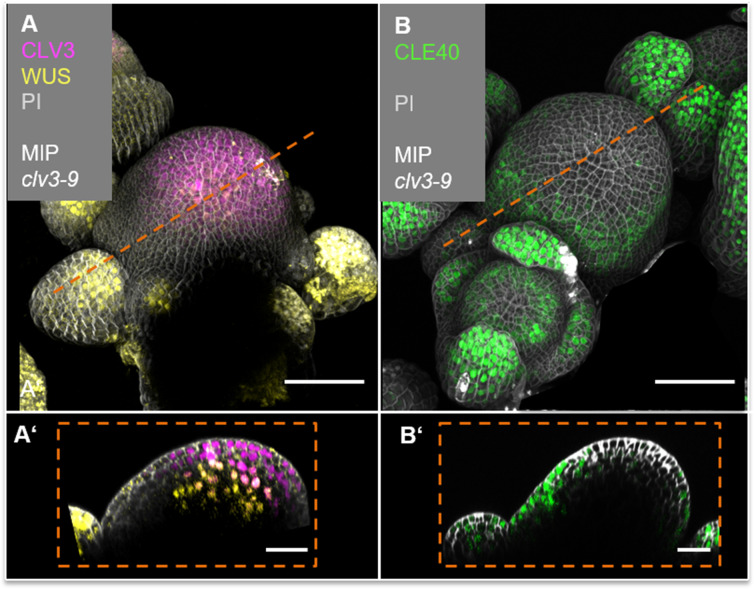
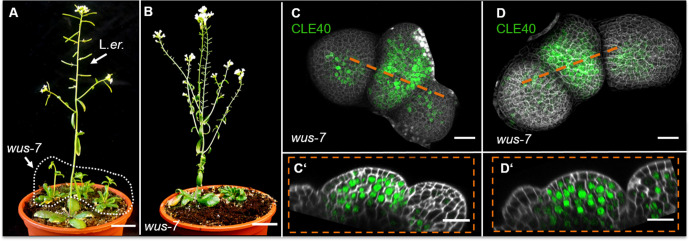
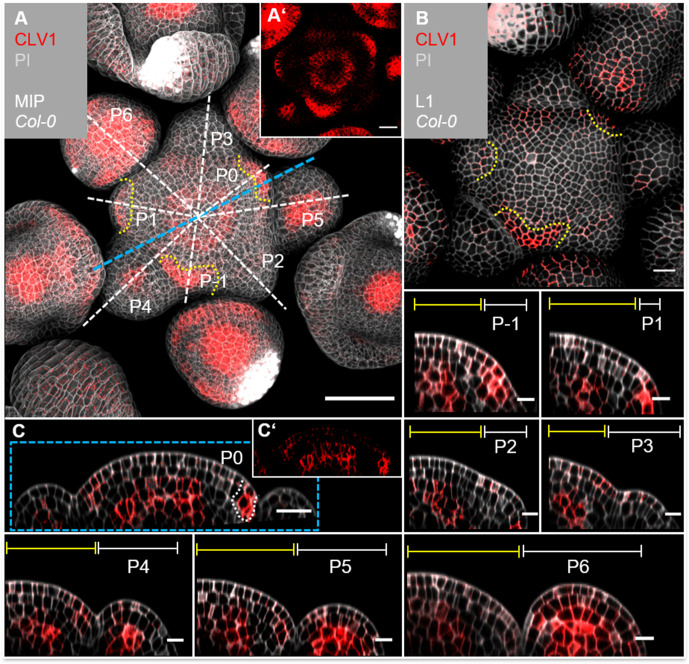
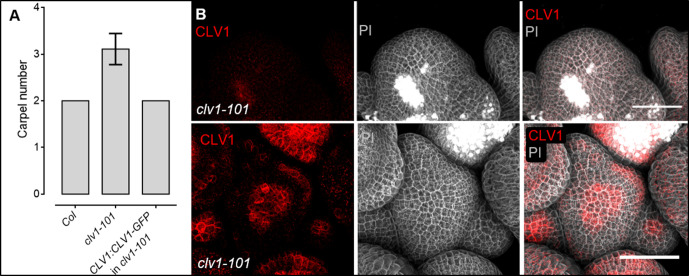
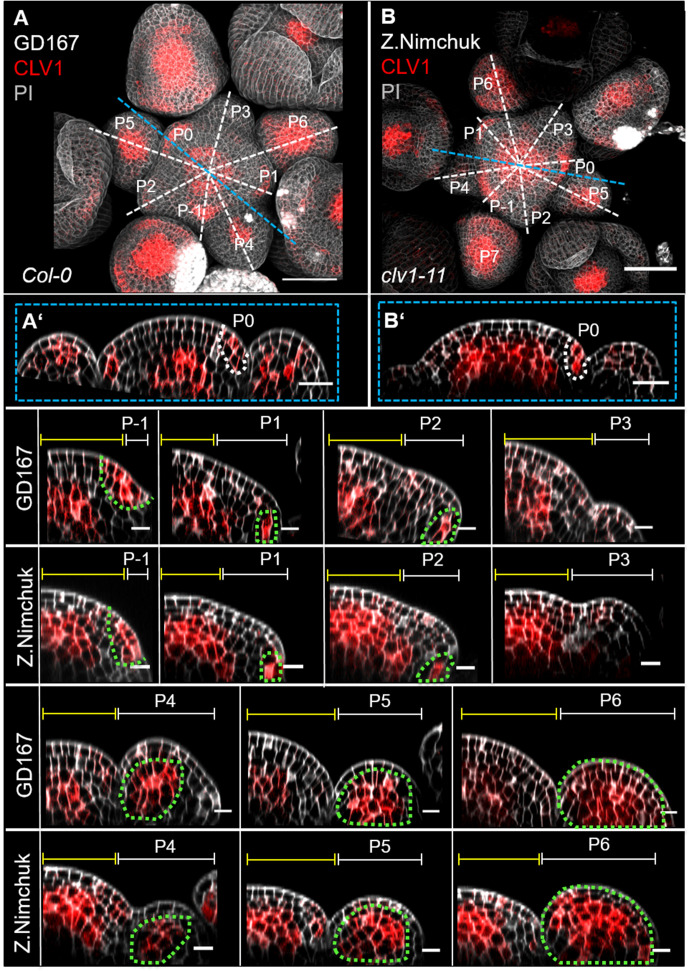
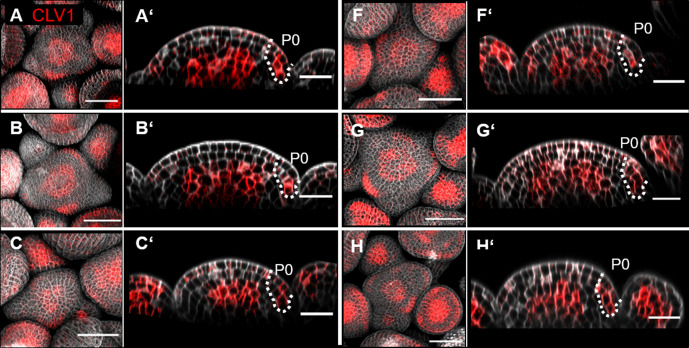
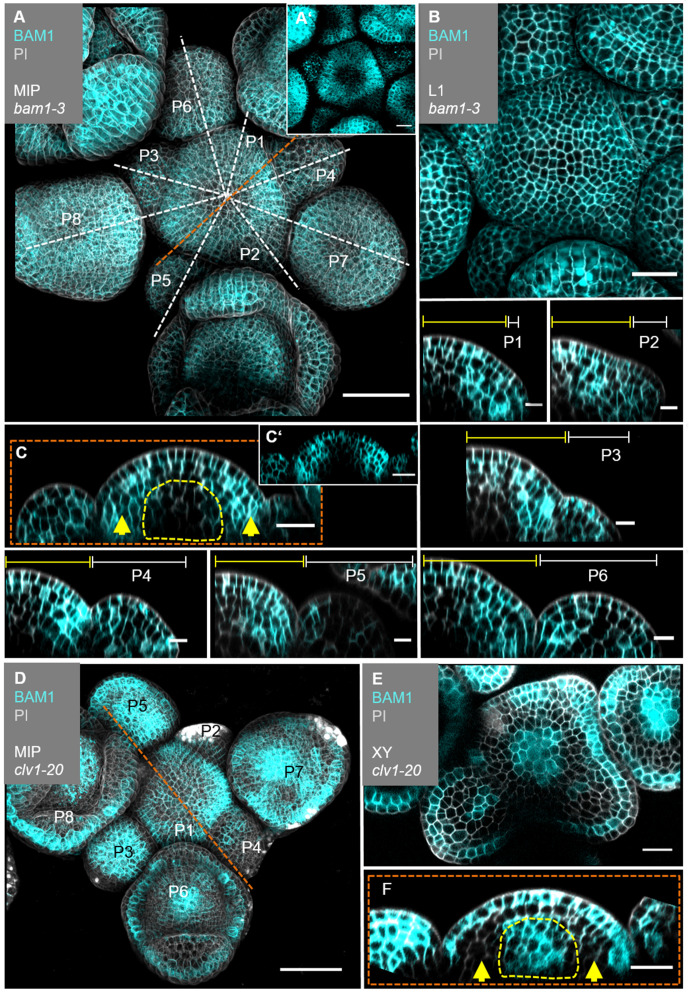
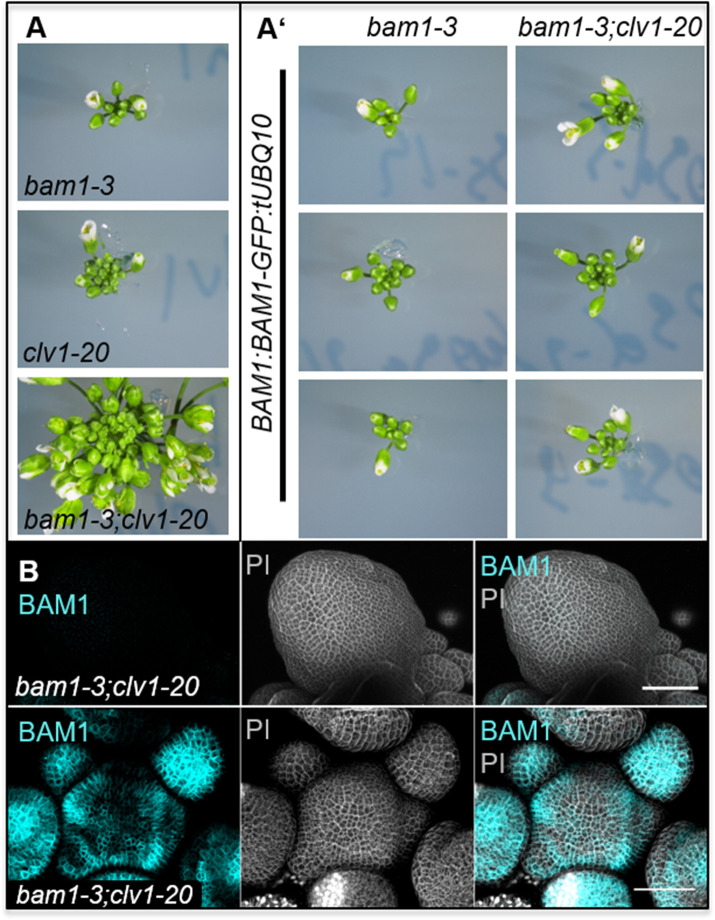
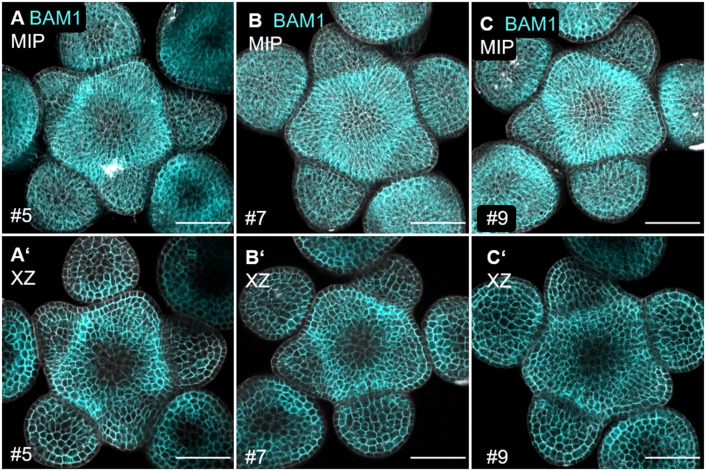
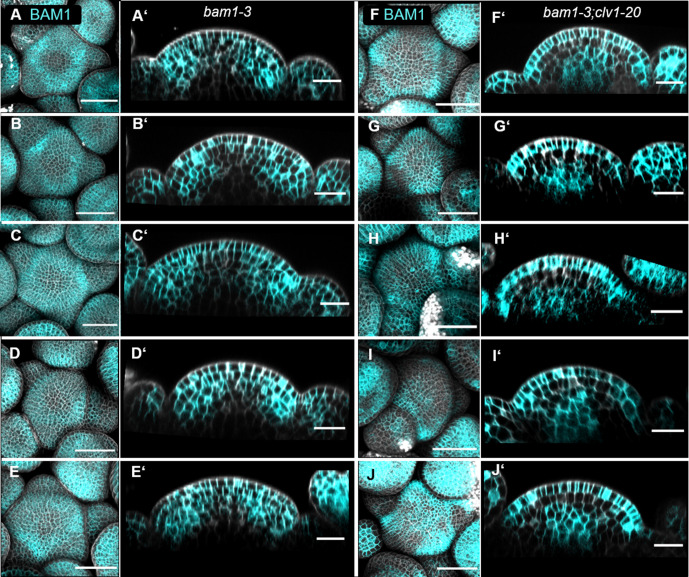
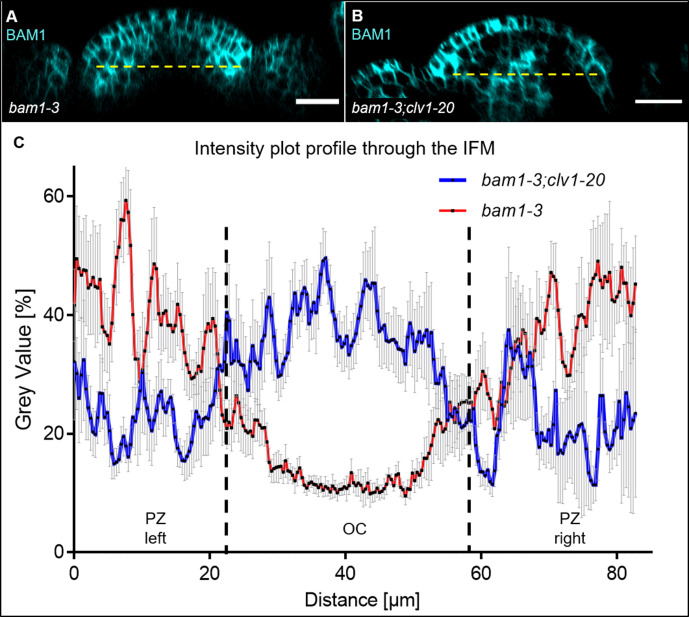
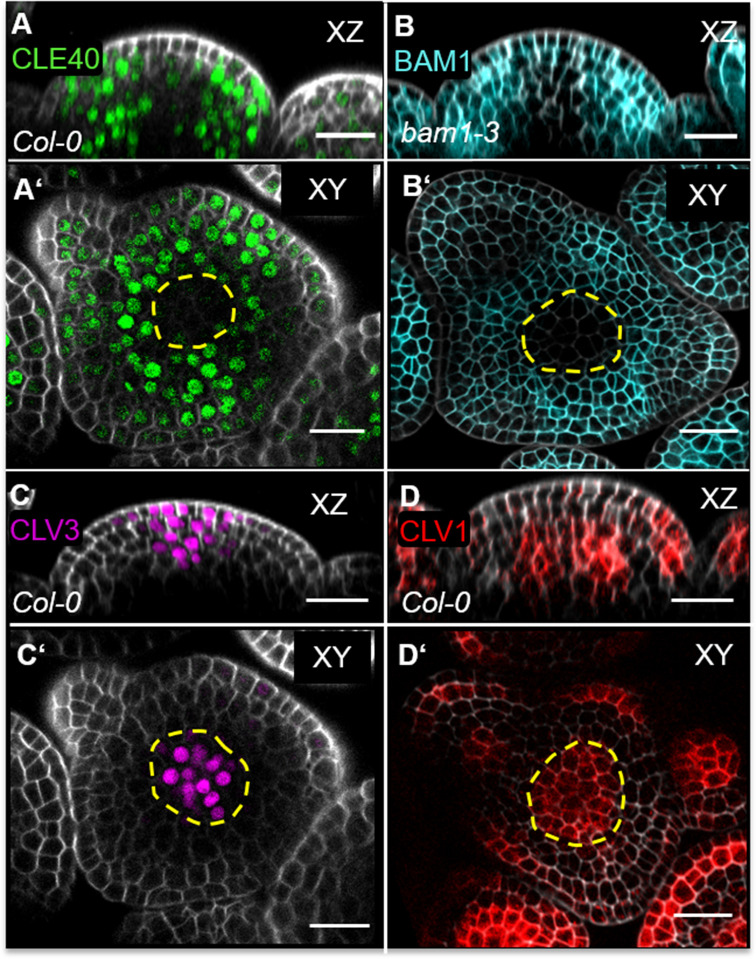
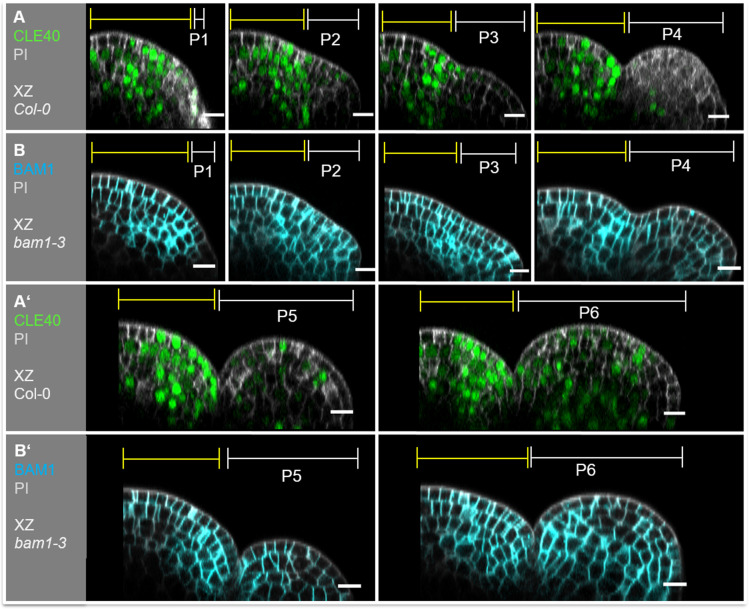
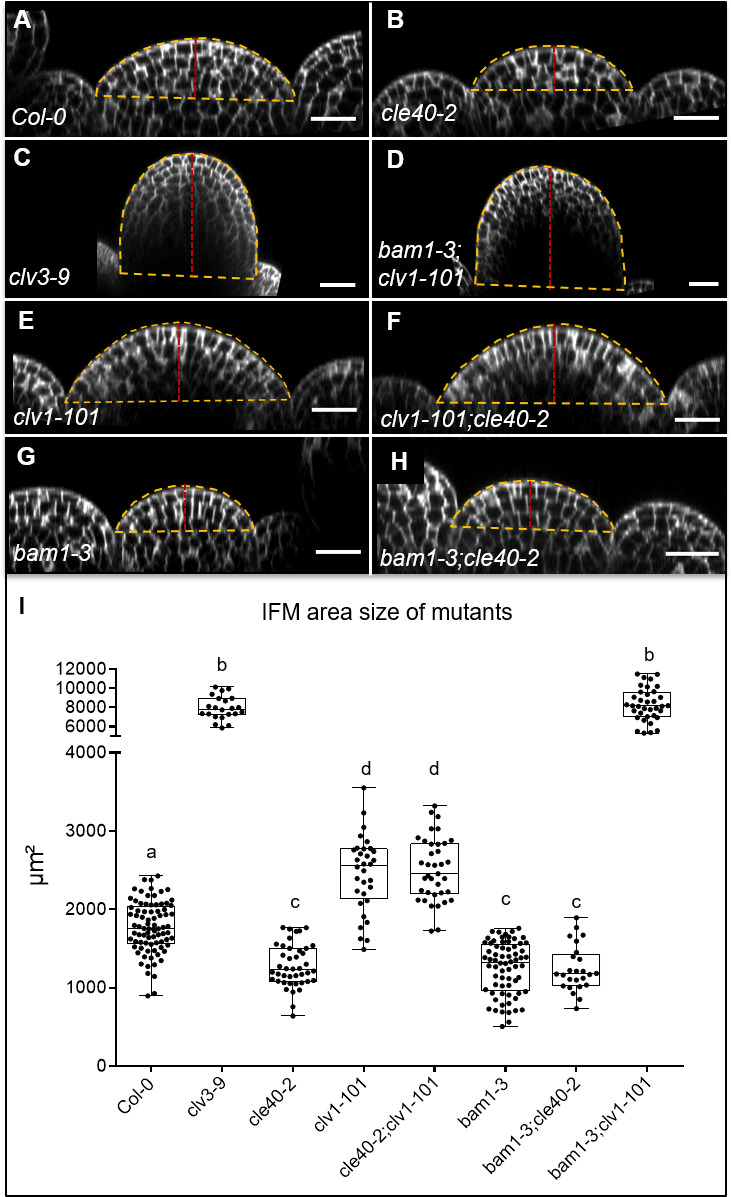
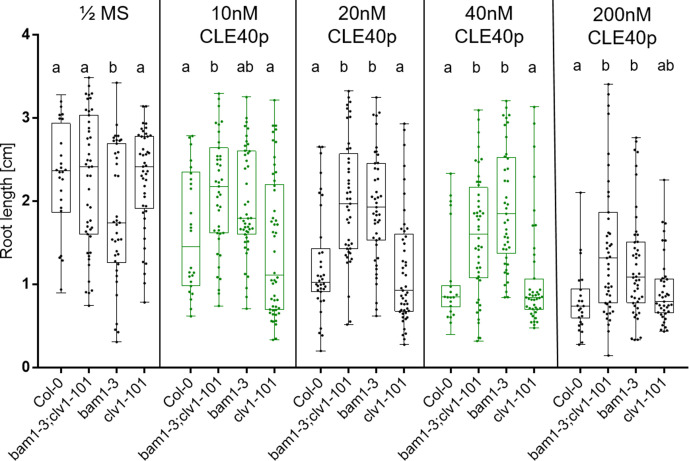
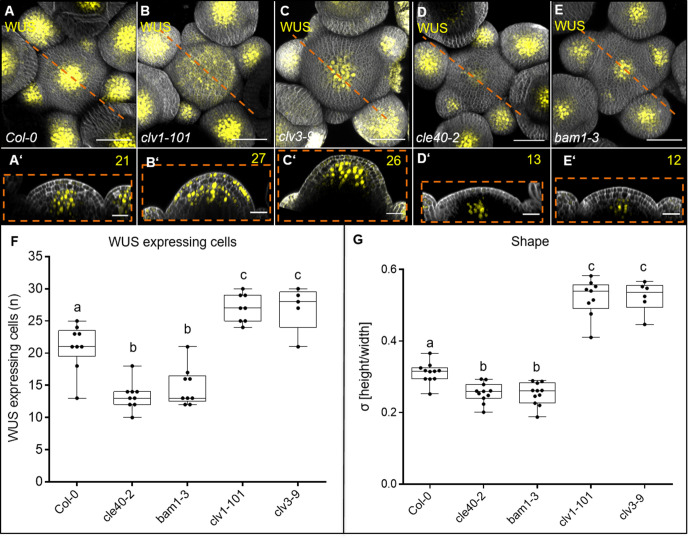
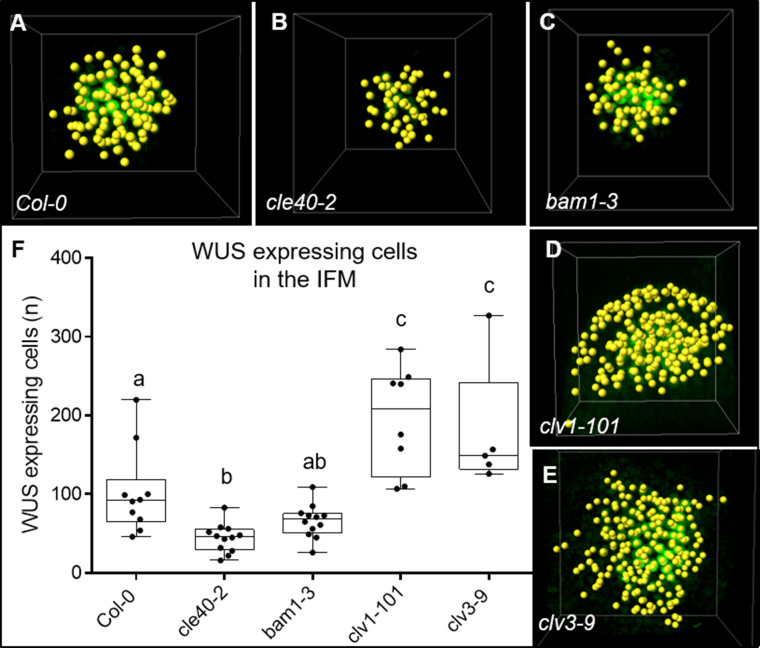
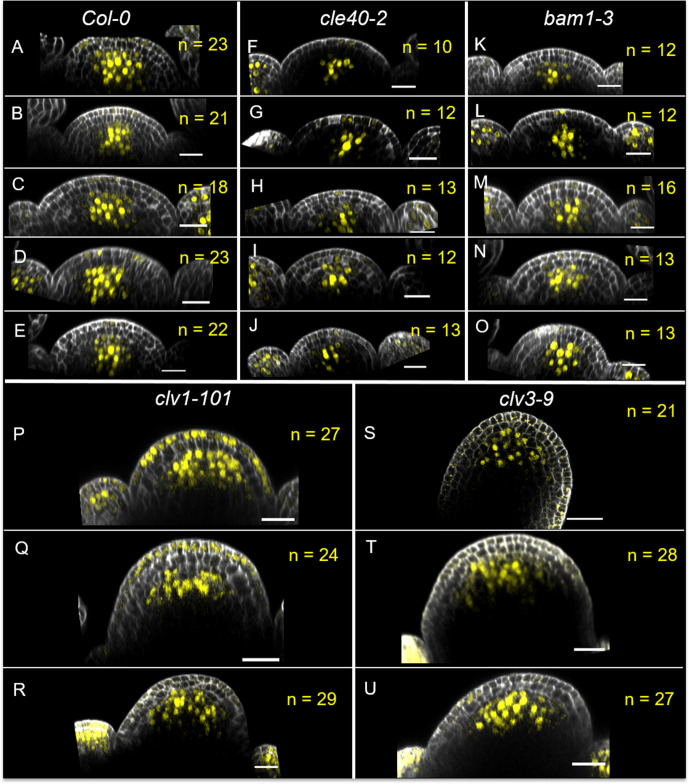
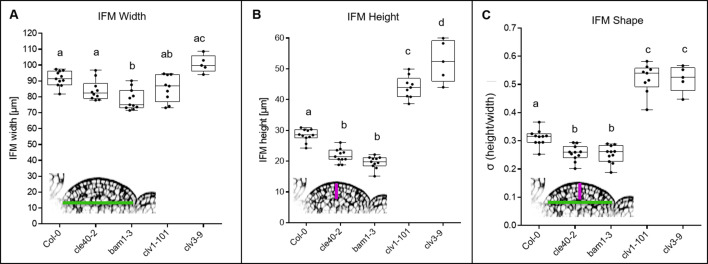
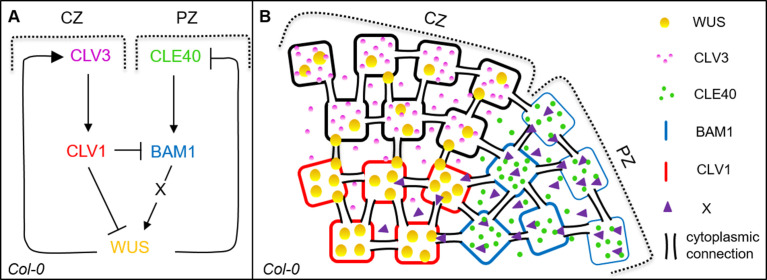
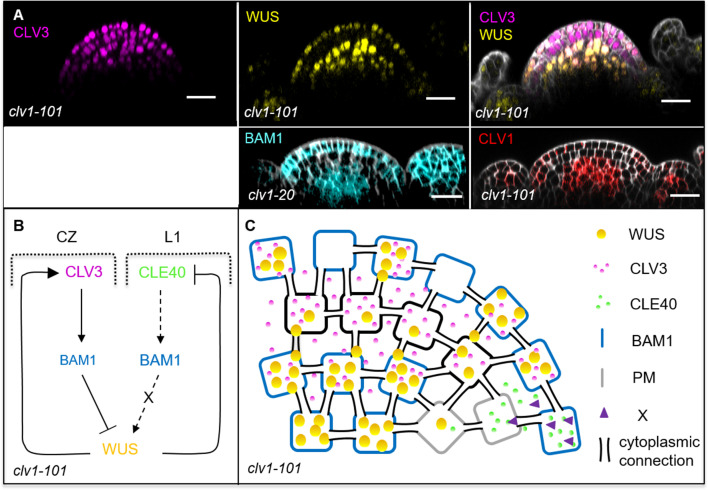
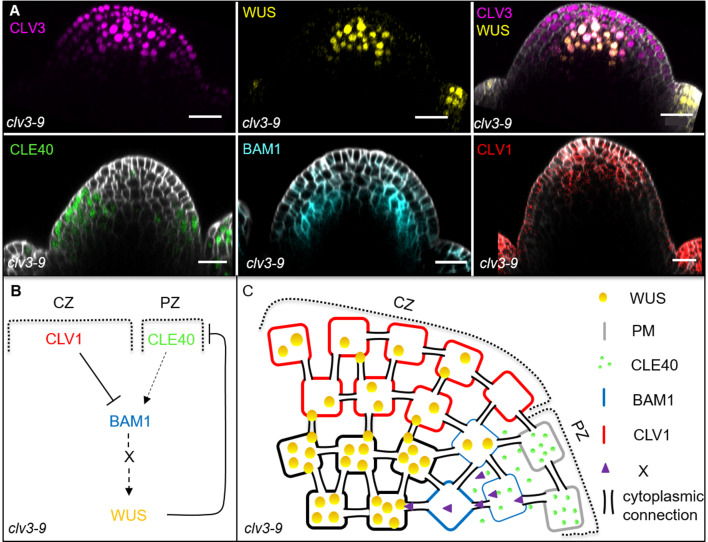
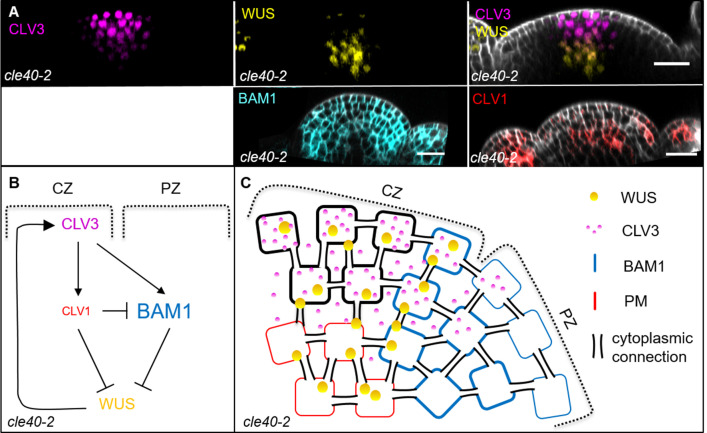
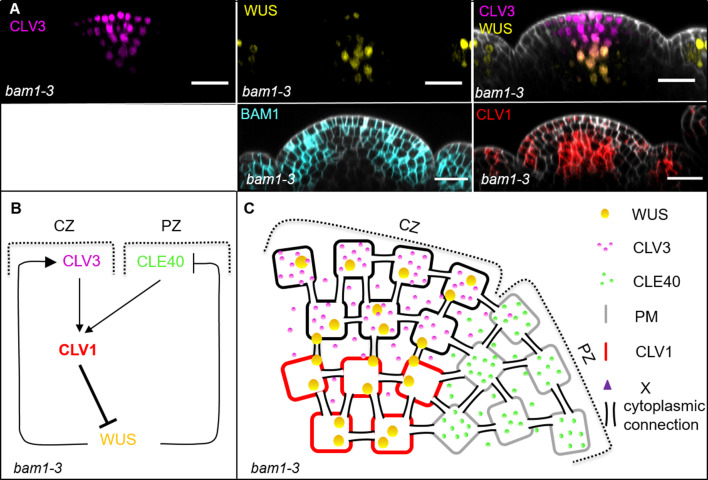
Stem cell homeostasis in plant shoot meristems requires tight coordination between stem cell proliferation and cell differentiation. In , stem cells express the secreted dodecapeptide CLAVATA3 (CLV3), which signals through the leucine-rich repeat (LRR)-receptor kinase CLAVATA1 (CLV1) and related CLV1-family members to downregulate expression of the homeodomain transcription factor (). WUS protein moves from cells below the stem cell domain to the meristem tip and promotes stem cell identity, together with expression, generating a negative feedback loop. How stem cell activity in the meristem centre is coordinated with organ initiation and cell differentiation at the periphery is unknown. We show here that the gene, encoding a secreted peptide closely related to CLV3, is expressed in the SAM in differentiating cells in a pattern complementary to that of promotes expression via BAM1, a CLV1-family receptor, and expression is in turn repressed in a -dependent manner. Together, establish a second negative feedback loop. We propose that stem cell homeostasis is achieved through two intertwined pathways that adjust WUS activity and incorporate information on the size of the stem cell domain, via , and on cell differentiation via .
植物茎尖分生组织中的干细胞稳态需要干细胞增殖和细胞分化之间的紧密协调。在这里,干细胞表达分泌的十二肽 CLAVATA3(CLV3),通过富含亮氨酸重复(LRR)-受体激酶 CLAVATA1(CLV1)和相关的 CLV1 家族成员信号传导,下调同源域转录因子()的表达。WUS 蛋白从干细胞域下方的细胞移动到分生组织顶端,并与表达一起促进干细胞特性,产生负反馈回路。干细胞活性如何在分生组织中心与外围的器官起始和细胞分化相协调尚不清楚。我们在这里表明,基因,编码一种与 CLV3 密切相关的分泌肽,在分化细胞中的 SAM 中以与表达模式互补的方式表达,通过 CLV1 家族受体 BAM1 促进表达,并且表达以的依赖性方式被抑制。共同作用,建立了第二个负反馈回路。我们提出,干细胞稳态是通过两条相互交织的途径实现的,通过调节 WUS 活性并通过整合干细胞域大小的信息(通过),以及通过细胞分化(通过)来实现的。