Department of Biology, New York University, New York, NY 10003, USA.
Department of Biology, New York University, New York, NY 10003, USA; New York Genome Center, New York, NY 10013, USA.
Cell Rep. 2022 Mar 15;38(11):110524. doi: 10.1016/j.celrep.2022.110524.
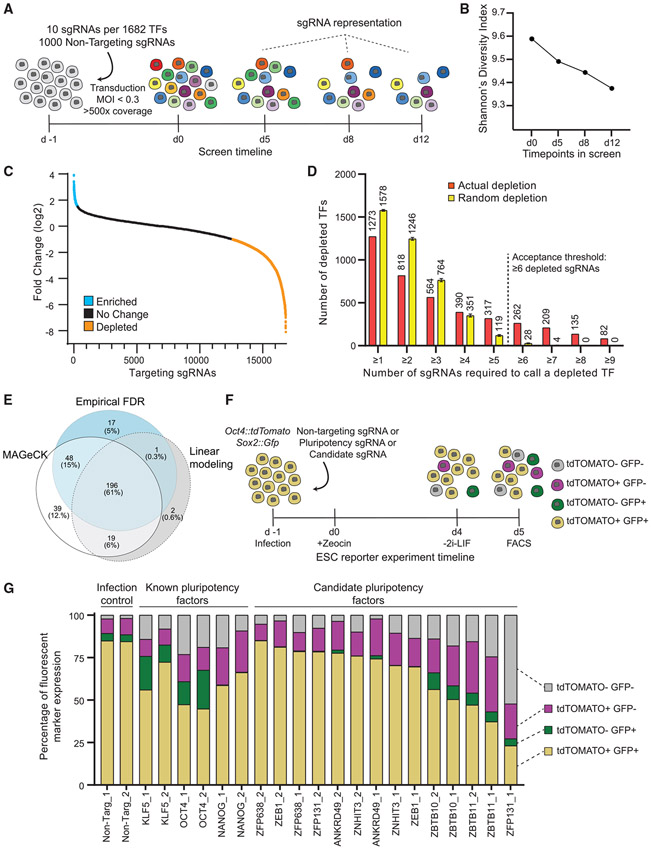
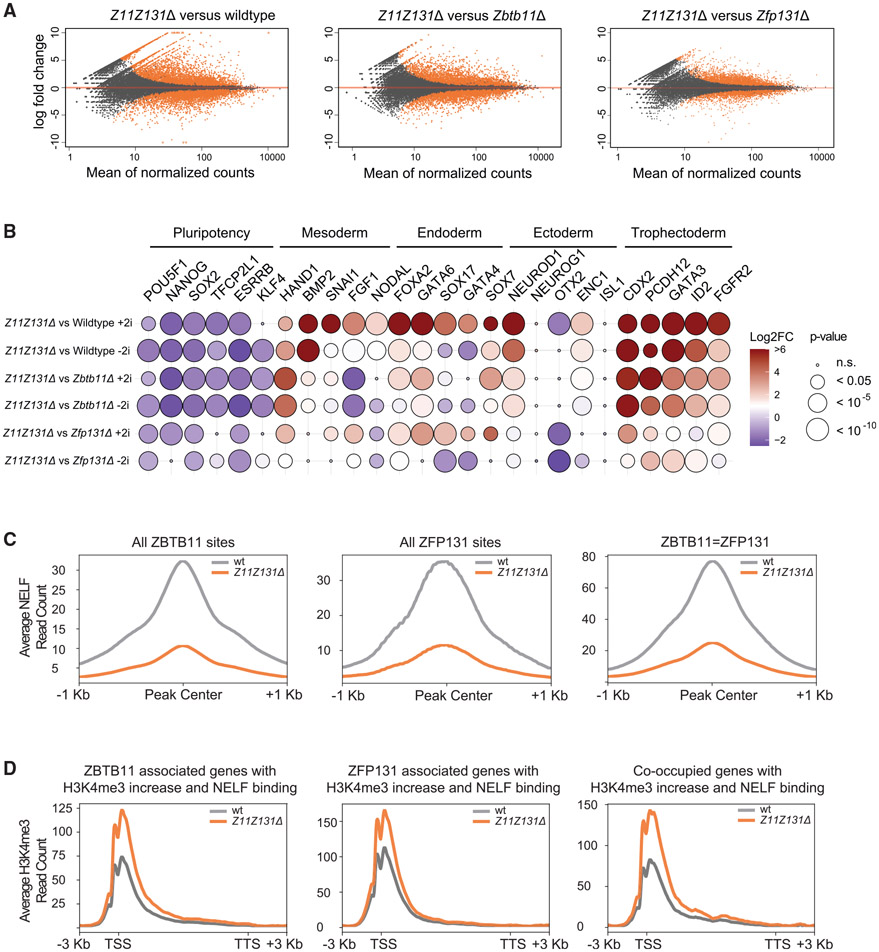
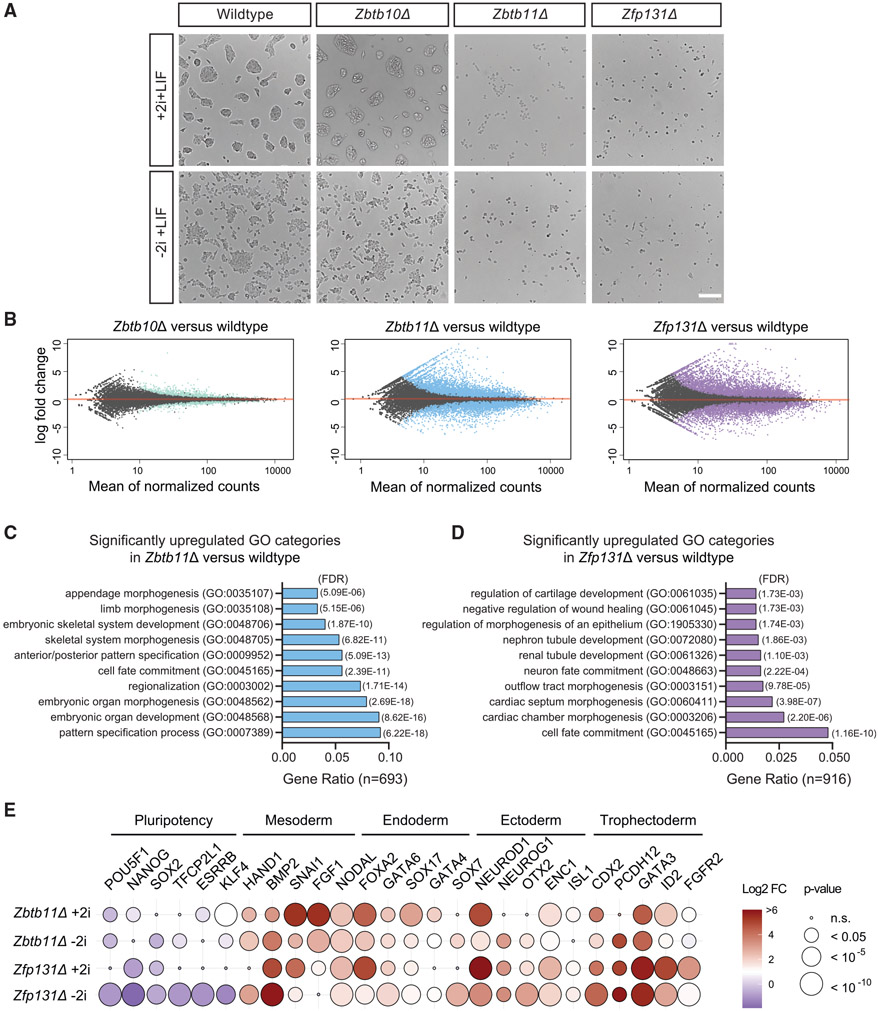
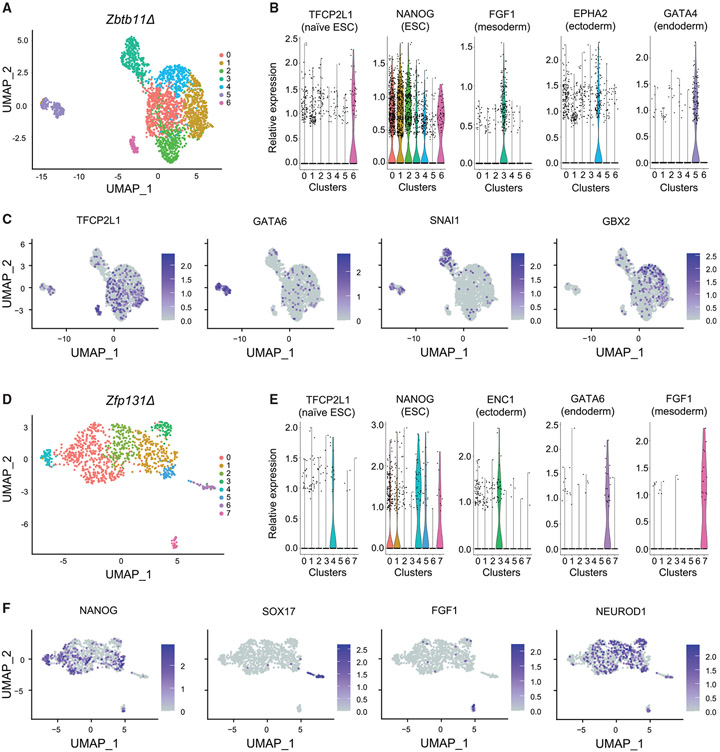
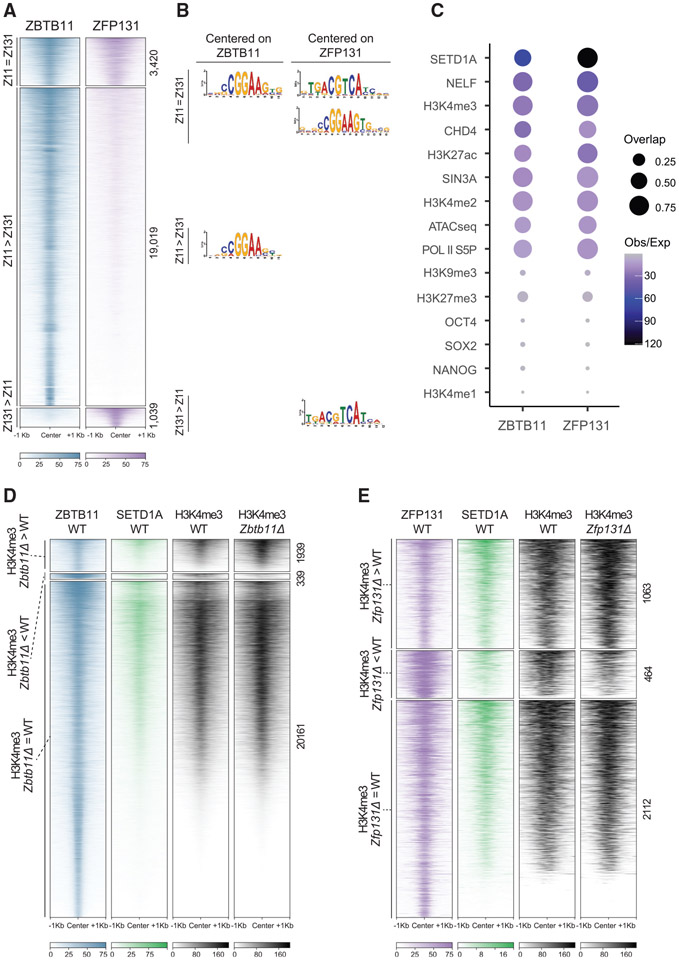
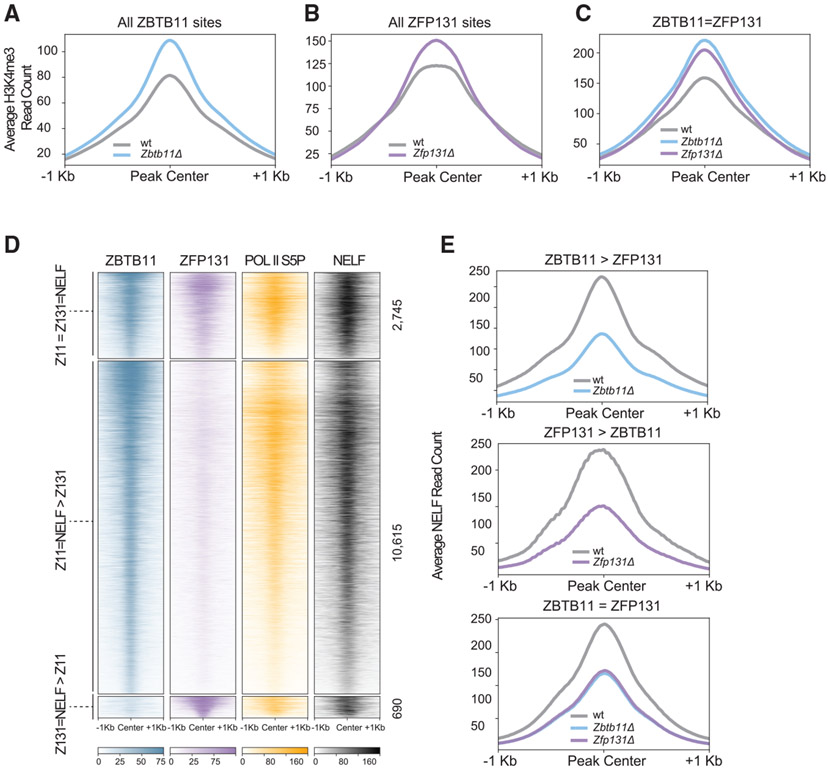
In pluripotent cells, a delicate activation-repression balance maintains pro-differentiation genes ready for rapid activation. The identity of transcription factors (TFs) that specifically repress pro-differentiation genes remains obscure. By targeting ∼1,700 TFs with CRISPR loss-of-function screen, we found that ZBTB11 and ZFP131 are required for embryonic stem cell (ESC) pluripotency. ESCs without ZBTB11 or ZFP131 lose colony morphology, reduce proliferation rate, and upregulate transcription of genes associated with three germ layers. ZBTB11 and ZFP131 bind proximally to pro-differentiation genes. ZBTB11 or ZFP131 loss leads to an increase in H3K4me3, negative elongation factor (NELF) complex release, and concomitant transcription at associated genes. Together, our results suggest that ZBTB11 and ZFP131 maintain pluripotency by preventing premature expression of pro-differentiation genes and present a generalizable framework to maintain cellular potency.
在多能细胞中,精细的激活-抑制平衡维持着促进分化的基因,使其能够快速激活。特异性抑制促分化基因的转录因子(TFs)的身份仍然不清楚。通过针对约 1700 个 TF 进行 CRISPR 功能丧失筛选,我们发现 ZBTB11 和 ZFP131 是胚胎干细胞(ESC)多能性所必需的。没有 ZBTB11 或 ZFP131 的 ESC 失去集落形态,降低增殖率,并上调与三个胚层相关的基因的转录。ZBTB11 和 ZFP131 与促分化基因近距离结合。ZBTB11 或 ZFP131 的缺失导致 H3K4me3 增加、负延伸因子(NELF)复合物释放以及相关基因的伴随转录。总之,我们的结果表明,ZBTB11 和 ZFP131 通过防止促分化基因的过早表达来维持多能性,并为维持细胞潜能提供了一种可推广的框架。