Centro de Investigación Biomédica en Red de Enfermedades Raras (CIBERER), Barcelona, Spain.
Universitat Pompeu Fabra (UPF), Barcelona, Spain.
Elife. 2022 Mar 18;11:e65310. doi: 10.7554/eLife.65310.
The identification of expression quantitative trait methylation (eQTMs), defined as associations between DNA methylation levels and gene expression, might help the biological interpretation of epigenome-wide association studies (EWAS). We aimed to identify autosomal cis eQTMs in children's blood, using data from 832 children of the Human Early Life Exposome (HELIX) project.
Blood DNA methylation and gene expression were measured with the Illumina 450K and the Affymetrix HTA v2 arrays, respectively. The relationship between methylation levels and expression of nearby genes (1 Mb window centered at the transcription start site, TSS) was assessed by fitting 13.6 M linear regressions adjusting for sex, age, cohort, and blood cell composition.
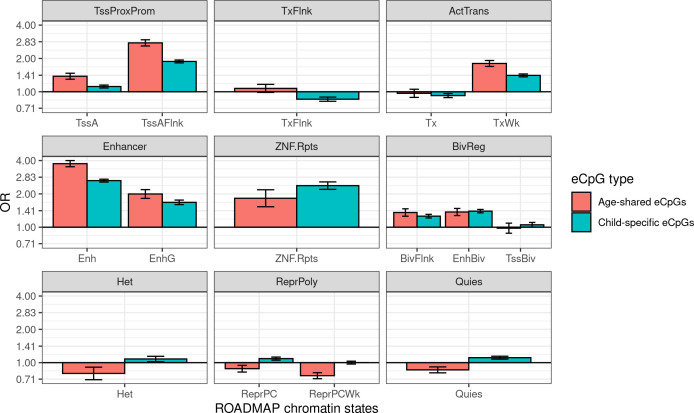
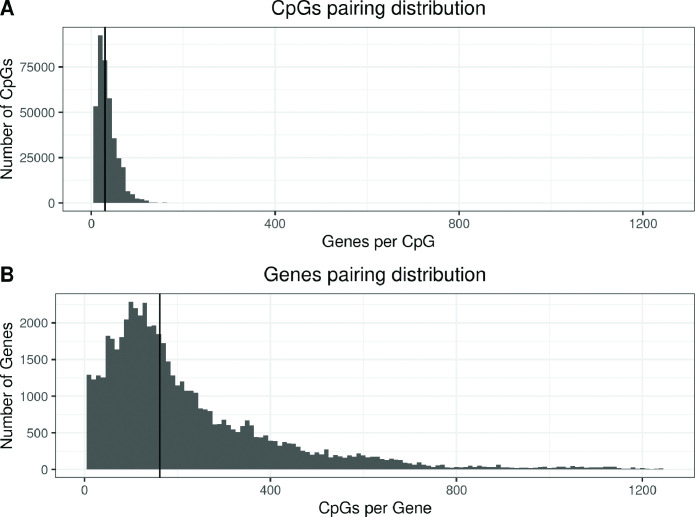
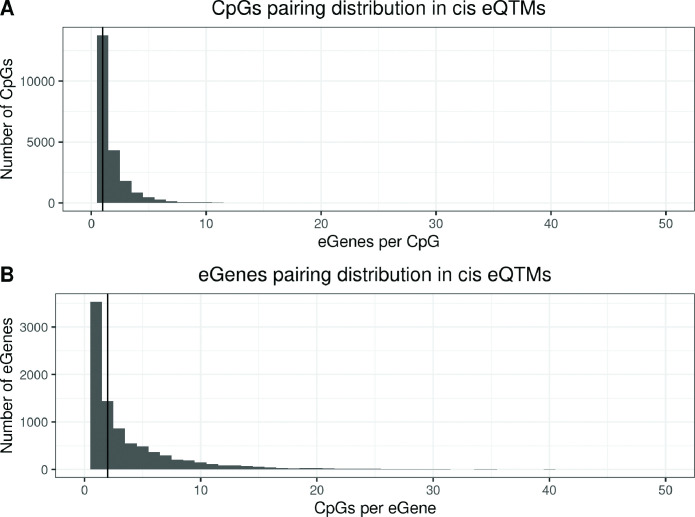
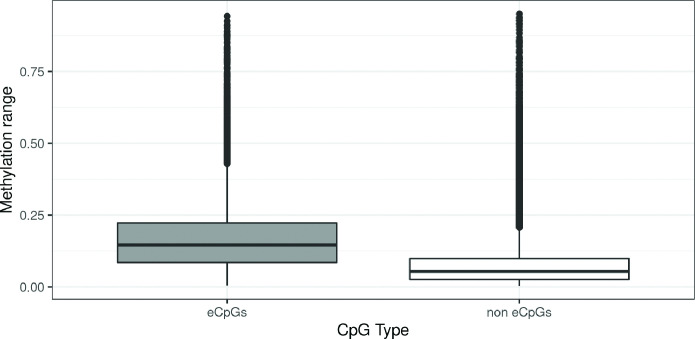
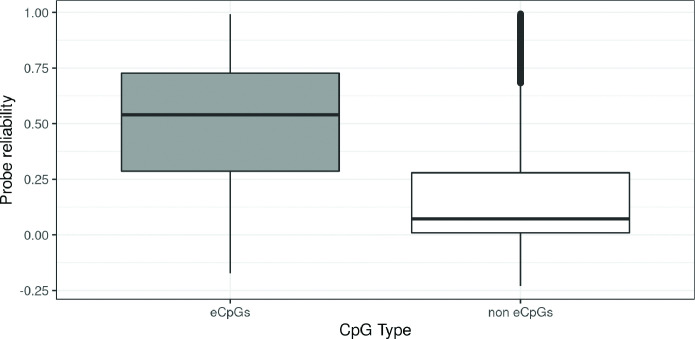
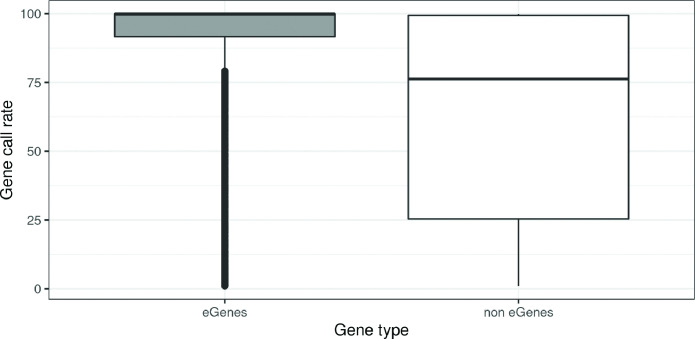
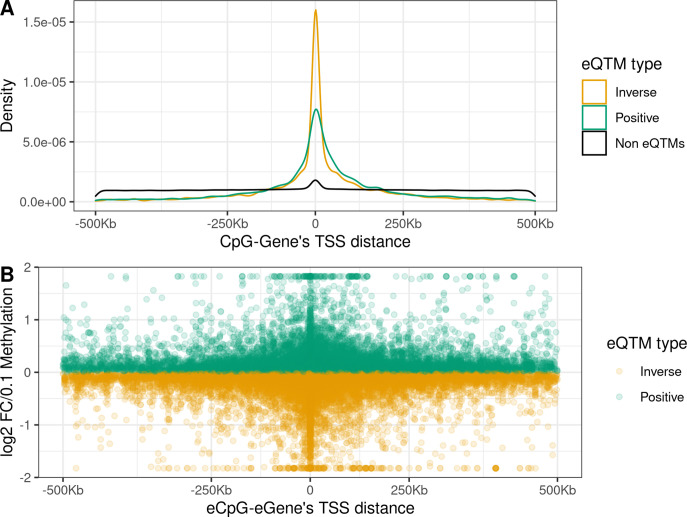
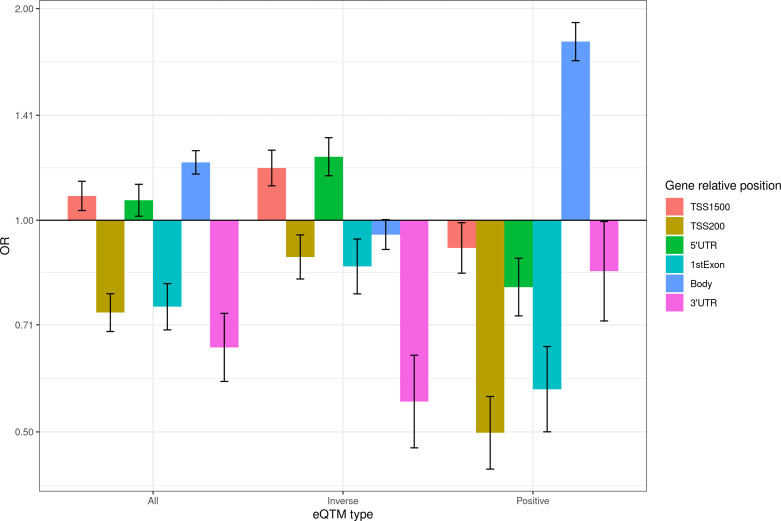
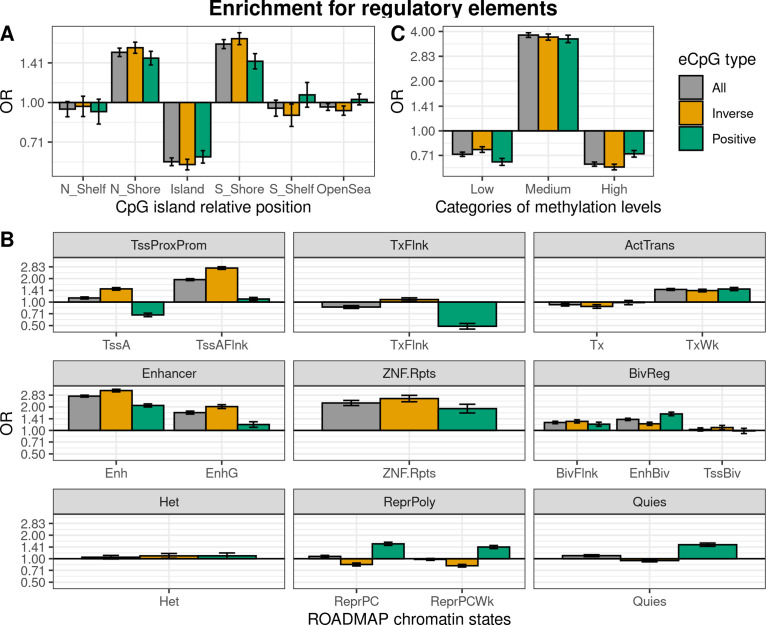
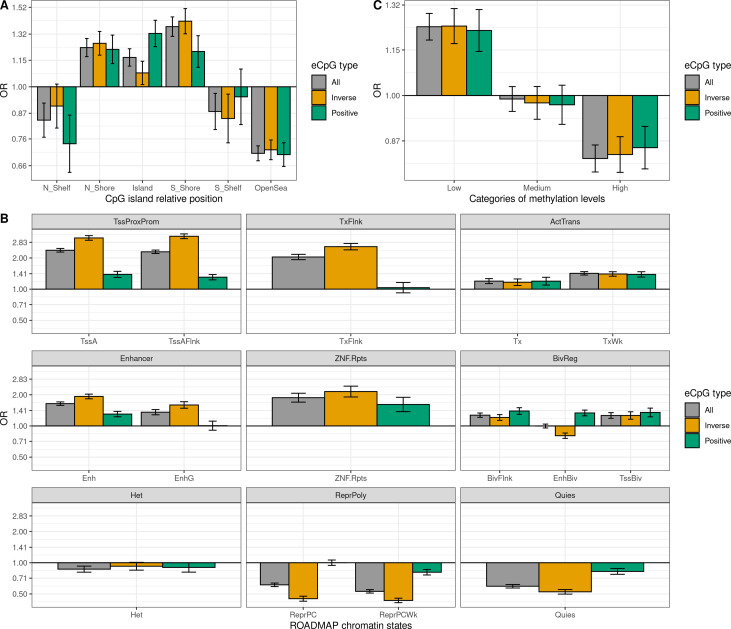
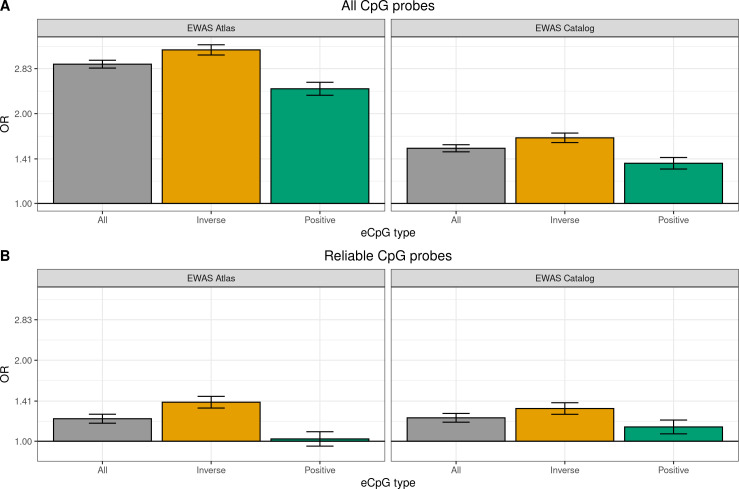
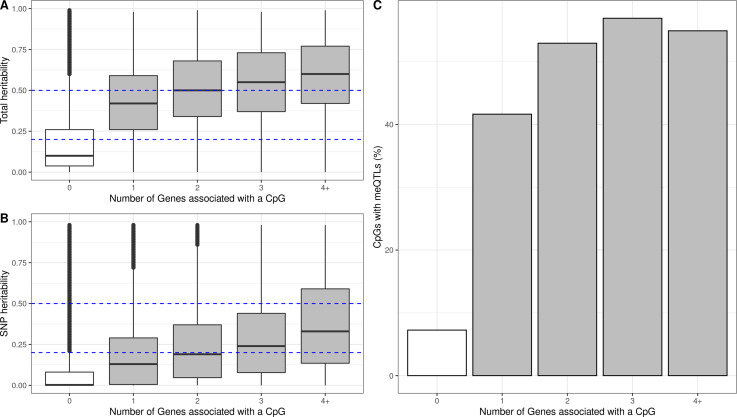
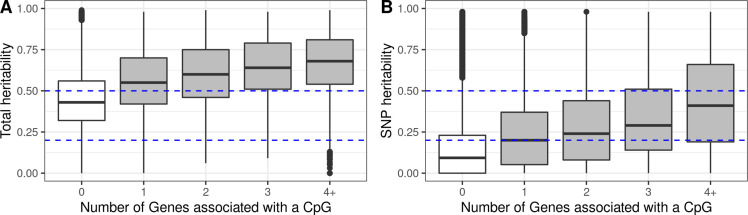
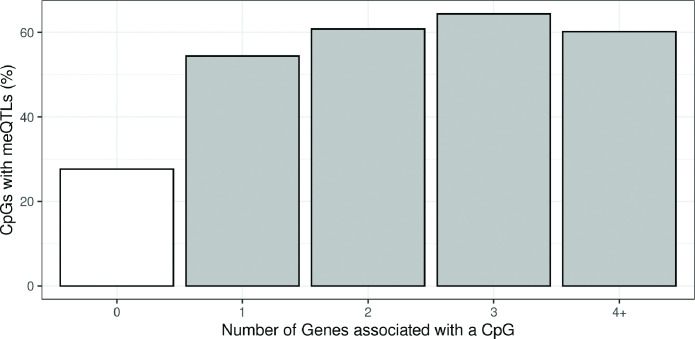
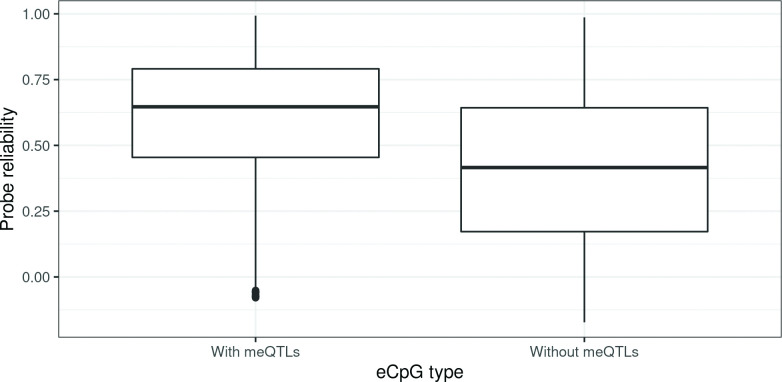
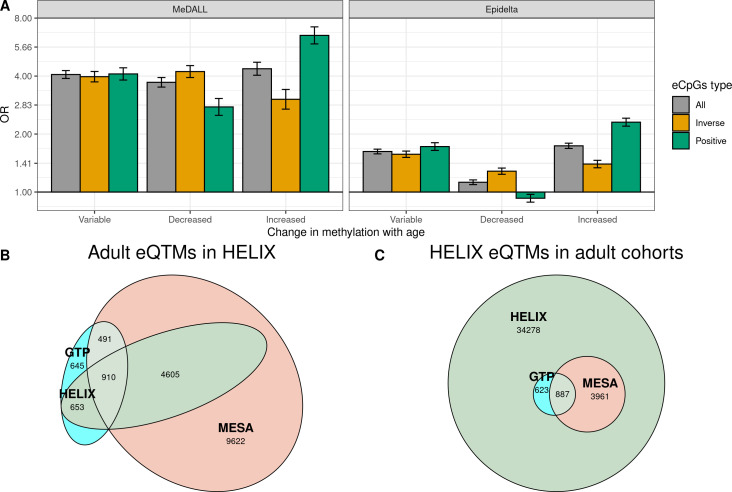
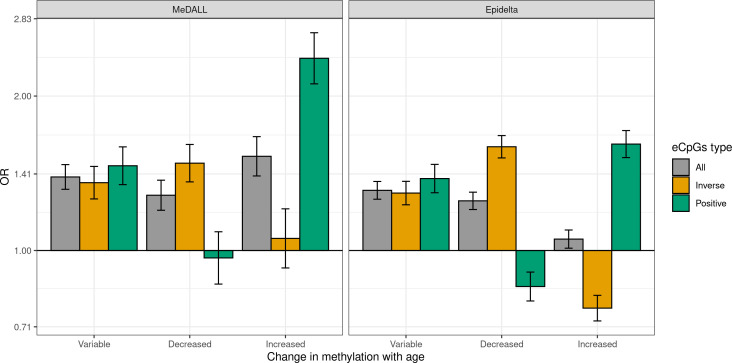
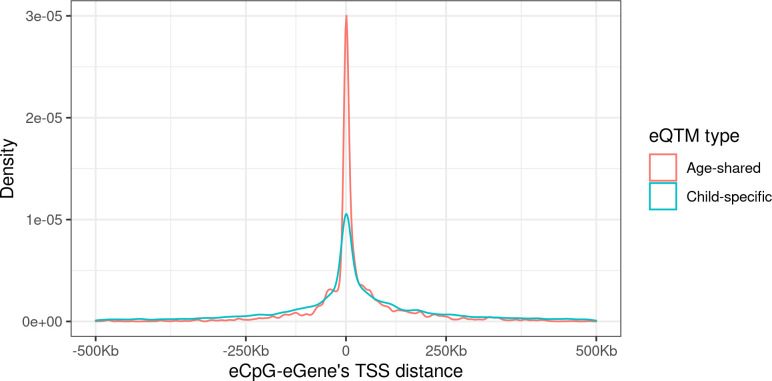
We identified 39,749 blood autosomal cis eQTMs, representing 21,966 unique CpGs (eCpGs, 5.7% of total CpGs) and 8,886 unique transcript clusters (eGenes, 15.3% of total transcript clusters, equivalent to genes). In 87.9% of these cis eQTMs, the eCpG was located at <250 kb from eGene's TSS; and 58.8% of all eQTMs showed an inverse relationship between the methylation and expression levels. Only around half of the autosomal cis-eQTMs eGenes could be captured through annotation of the eCpG to the closest gene. eCpGs had less measurement error and were enriched for active blood regulatory regions and for CpGs reported to be associated with environmental exposures or phenotypic traits. In 40.4% of the eQTMs, the CpG and the eGene were both associated with at least one genetic variant. The overlap of autosomal cis eQTMs in children's blood with those described in adults was small (13.8%), and age-shared cis eQTMs tended to be proximal to the TSS and enriched for genetic variants.
This catalogue of autosomal cis eQTMs in children's blood can help the biological interpretation of EWAS findings and is publicly available at https://helixomics.isglobal.org/ and at Dryad (doi:10.5061/dryad.fxpnvx0t0).
The study has received funding from the European Community's Seventh Framework Programme (FP7/2007-206) under grant agreement no 308333 (HELIX project); the H2020-EU.3.1.2. - Preventing Disease Programme under grant agreement no 874583 (ATHLETE project); from the European Union's Horizon 2020 research and innovation programme under grant agreement no 733206 (LIFECYCLE project), and from the European Joint Programming Initiative "A Healthy Diet for a Healthy Life" (JPI HDHL and Instituto de Salud Carlos III) under the grant agreement no AC18/00006 (NutriPROGRAM project). The genotyping was supported by the projects PI17/01225 and PI17/01935, funded by the Instituto de Salud Carlos III and co-funded by European Union (ERDF, "A way to make Europe") and the Centro Nacional de Genotipado-CEGEN (PRB2-ISCIII). BiB received core infrastructure funding from the Wellcome Trust (WT101597MA) and a joint grant from the UK Medical Research Council (MRC) and Economic and Social Science Research Council (ESRC) (MR/N024397/1). INMA data collections were supported by grants from the Instituto de Salud Carlos III, CIBERESP, and the Generalitat de Catalunya-CIRIT. KANC was funded by the grant of the Lithuanian Agency for Science Innovation and Technology (6-04-2014_31V-66). The Norwegian Mother, Father and Child Cohort Study is supported by the Norwegian Ministry of Health and Care Services and the Ministry of Education and Research. The Rhea project was financially supported by European projects (EU FP6-2003-Food-3-NewGeneris, EU FP6. STREP Hiwate, EU FP7 ENV.2007.1.2.2.2. Project No 211250 Escape, EU FP7-2008-ENV-1.2.1.4 Envirogenomarkers, EU FP7-HEALTH-2009- single stage CHICOS, EU FP7 ENV.2008.1.2.1.6. Proposal No 226285 ENRIECO, EU- FP7- HEALTH-2012 Proposal No 308333 HELIX), and the Greek Ministry of Health (Program of Prevention of obesity and neurodevelopmental disorders in preschool children, in Heraklion district, Crete, Greece: 2011-2014; "Rhea Plus": Primary Prevention Program of Environmental Risk Factors for Reproductive Health, and Child Health: 2012-15). We acknowledge support from the Spanish Ministry of Science and Innovation through the "Centro de Excelencia Severo Ochoa 2019-2023" Program (CEX2018-000806-S), and support from the Generalitat de Catalunya through the CERCA Program. MV-U and CR-A were supported by a FI fellowship from the Catalan Government (FI-DGR 2015 and #016FI_B 00272). MC received funding from Instituto Carlos III (Ministry of Economy and Competitiveness) (CD12/00563 and MS16/00128).

识别表达数量性状甲基化(eQTM),定义为 DNA 甲基化水平与基因表达之间的关联,可能有助于对全基因组关联研究(EWAS)的生物解释。我们旨在使用来自人类早期生活暴露组学(HELIX)项目的 832 名儿童的数据,确定儿童血液中的常染色体顺式 eQTM。
使用 Illumina 450K 和 Affymetrix HTA v2 芯片分别测量血液 DNA 甲基化和基因表达。通过拟合 13.6M 个线性回归来评估附近基因(转录起始位点 1Mb 窗口)的甲基化水平和表达之间的关系,这些回归通过性别、年龄、队列和血细胞组成进行调整。
我们鉴定了 39749 个血液常染色体顺式 eQTM,代表 21966 个独特的 CpG(占总 CpG 的 5.7%)和 8886 个独特的转录簇(eGenes,占总转录簇的 15.3%,相当于基因)。在这些顺式 eQTM 中,87.9%的 eCpG 位于 eGene 的 TSS 附近<250kb;并且 58.8%的所有 eQTM 显示出甲基化和表达水平之间的反比关系。只有大约一半的常染色体顺式-eQTM 的 eGenes 可以通过注释 eCpG 到最近的基因来捕获。eCpGs 的测量误差较小,并且富含活跃的血液调控区域,以及与环境暴露或表型特征相关的报道 CpG。在 40.4%的 eQTM 中,CpG 和 eGene 都与至少一个遗传变异有关。儿童血液中的常染色体顺式 eQTM 与成人中描述的 eQTM 重叠很小(13.8%),并且年龄共享的顺式 eQTM 倾向于靠近 TSS,并富含遗传变异。
本研究目录中的儿童血液常染色体顺式 eQTM 可帮助对 EWAS 结果进行生物解释,该目录可在 https://helixomics.isglobal.org/ 和 Dryad(doi:10.5061/dryad.fxpnvx0t0)上获得。
该研究得到了欧盟第七框架计划(FP7/2007-2016)的资助(HELIX 项目);欧盟地平线 2020 研究与创新计划(ATHLETE 项目);欧盟地平线 2020 研究与创新计划(874583 号);以及欧洲联合规划倡议“健康生活的健康饮食”(JPI HDHL 和 Instituto de Salud Carlos III),资助号为 AC18/00006(NutriPROGRAM 项目)。基因分型由项目 PI17/01225 和 PI17/01935 提供支持,这些项目由西班牙卡洛斯三世健康研究所(Instituto de Salud Carlos III)和欧盟共同资助(ERDF,“走向欧洲”)和国家基因分型中心-CEGEN(PRB2-ISCIII)。BiB 得到了威康信托基金会(Wellcome Trust)核心基础设施基金(WT101597MA)和英国医学研究理事会(MRC)和经济和社会研究理事会(ESRC)联合资助的联合资助。INMA 数据收集得到了西班牙卡洛斯三世健康研究所、CIBERESP 和加泰罗尼亚自治区- CIRCIT 的资助。KANC 得到了立陶宛科学创新和技术署(6-04-2014_31V-66)的拨款。挪威母亲、父亲和儿童队列研究由挪威卫生部和卫生部以及教育部和研究部资助。Rhea 项目得到了欧盟项目(欧盟第六框架计划-2003 年-Food-3-NewGeneris,欧盟第六框架计划-STRP Hiwate,欧盟第七框架计划 ENV.2007.1.2.2.2.项目号 211250 Escape,欧盟第七框架计划-ENV.2008.1.2.1.2.项目号 226285 Envirogenomarkers,欧盟第七框架计划-HEALTH-2009-单一阶段 CHICOS,欧盟第七框架计划-ENV.2008.1.2.1.6.提案号 226285 ENRIECO,欧盟第七框架计划-HEALTH-2012 号提案 308333 HELIX)和希腊卫生部(在 Heraklion 区预防学龄前儿童肥胖和神经发育障碍的计划,2011-2014 年;“Rhea Plus”:生殖健康和儿童健康的环境风险因素初级预防计划,2012-15 年)的资助。我们感谢西班牙科学创新部通过“Severo Ochoa 卓越中心 2019-2023”计划(CEX2018-000806-S)提供的支持,以及加泰罗尼亚自治区通过 CERCA 计划提供的支持。MV-U 和 CR-A 得到了加泰罗尼亚政府 FI 奖学金的支持(FI-DGR 2015 和 #016FI_B 00272)。MC 得到了西班牙卡洛斯三世健康研究所(西班牙经济和竞争力部)(CD12/00563 和 MS16/00128)的资助。