Spietz Rachel L, Payne Devon, Kulkarni Gargi, Metcalf William W, Roden Eric E, Boyd Eric S
Department of Microbiology and Cell Biology, Montana State University, Bozeman, MT, United States.
Department of Microbiology, University of Illinois at Urbana-Champaign, Urbana, IL, United States.
Front Microbiol. 2022 May 9;13:878387. doi: 10.3389/fmicb.2022.878387. eCollection 2022.
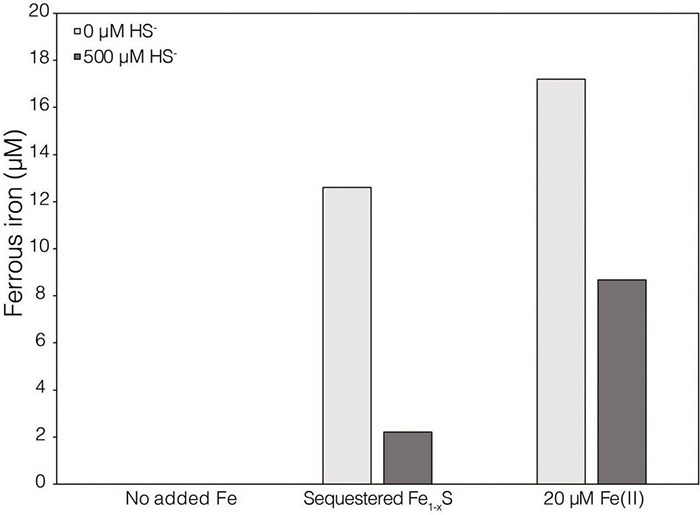
Pyrite (FeS) has a very low solubility and therefore has historically been considered a sink for iron (Fe) and sulfur (S) and unavailable to biology in the absence of oxygen and oxidative weathering. Anaerobic methanogens were recently shown to reduce FeS and assimilate Fe and S reduction products to meet nutrient demands. However, the mechanism of FeS mineral reduction and the forms of Fe and S assimilated by methanogens remained unclear. Thermodynamic calculations described herein indicate that H at aqueous concentrations as low as 10 M favors the reduction of FeS, with sulfide (HS) and pyrrhotite (Fe S) as products; abiotic laboratory experiments confirmed the reduction of FeS with dissolved H concentrations greater than 1.98 × 10 M H. Growth studies of provided with FeS as the sole source of Fe and S resulted in H production but at concentrations too low to drive abiotic FeS reduction, based on abiotic laboratory experimental data. A strain of with deletions in all [NiFe]-hydrogenases maintained the ability to reduce FeS during growth, providing further evidence that extracellular electron transport (EET) to FeS does not involve H or [NiFe]-hydrogenases. Physical contact between cells and FeS was required for mineral reduction but was not required to obtain Fe and S from dissolution products. The addition of a synthetic electron shuttle, anthraquinone-2,6-disulfonate, allowed for biological reduction of FeS when physical contact between cells and FeS was prohibited, indicating that exogenous electron shuttles can mediate FeS reduction. Transcriptomics experiments revealed upregulation of several cytoplasmic oxidoreductases during growth of on FeS, which may indicate involvement in provisioning low potential electrons for EET to FeS. Collectively, the data presented herein indicate that reduction of insoluble FeS by occurred electron transfer from the cell surface to the mineral surface resulting in the generation of soluble HS and mineral-associated Fe S. Solubilized Fe(II), but not HS, from mineral-associated Fe S reacts with aqueous HS yielding aqueous iron sulfur clusters (FeS ) that likely serve as the Fe and S source for methanogen growth and activity. FeS nucleation and subsequent precipitation on the surface of cells may result in accelerated EET to FeS, resulting in positive feedback between cell activity and FeS reduction.
黄铁矿(FeS)的溶解度极低,因此在历史上一直被视为铁(Fe)和硫(S)的汇,在无氧和氧化风化的情况下生物无法利用。最近有研究表明,厌氧产甲烷菌能够还原FeS,并同化Fe和S的还原产物以满足营养需求。然而,FeS矿物还原的机制以及产甲烷菌同化的Fe和S的形式仍不清楚。本文所述的热力学计算表明,水溶液中低至10 M的H有利于FeS的还原,产物为硫化物(HS)和磁黄铁矿(Fe S);非生物实验室实验证实,当溶解的H浓度大于1.98×10 M H时,FeS会发生还原。以FeS作为唯一Fe和S来源的生长研究产生了H,但根据非生物实验室实验数据,其浓度过低,无法驱动非生物FeS还原。一株所有[NiFe] - 氢化酶均缺失的菌株在生长过程中仍保持还原FeS的能力,这进一步证明向FeS的细胞外电子传递(EET)不涉及H或[NiFe] - 氢化酶。矿物还原需要细胞与FeS之间的物理接触,但从溶解产物中获取Fe和S则不需要。当细胞与FeS之间的物理接触被禁止时,添加合成电子穿梭体蒽醌 - 2,6 - 二磺酸盐可实现FeS的生物还原,这表明外源电子穿梭体可以介导FeS还原。转录组学实验表明,在以FeS为生长底物时,几种细胞质氧化还原酶会上调,这可能表明它们参与为向FeS的EET提供低电位电子。总体而言,本文提供的数据表明,通过细胞表面向矿物表面的电子转移发生了对不溶性FeS的还原,从而产生了可溶性HS和与矿物相关的Fe S。与矿物相关的Fe S中溶解的Fe(II)而非HS与水溶液中的HS反应,生成水溶液中的铁硫簇(FeS ),这些簇可能作为产甲烷菌生长和活性的Fe和S来源。FeS 在细胞表面的成核及随后的沉淀可能导致向FeS的EET加速,从而在细胞活性和FeS还原之间产生正反馈。