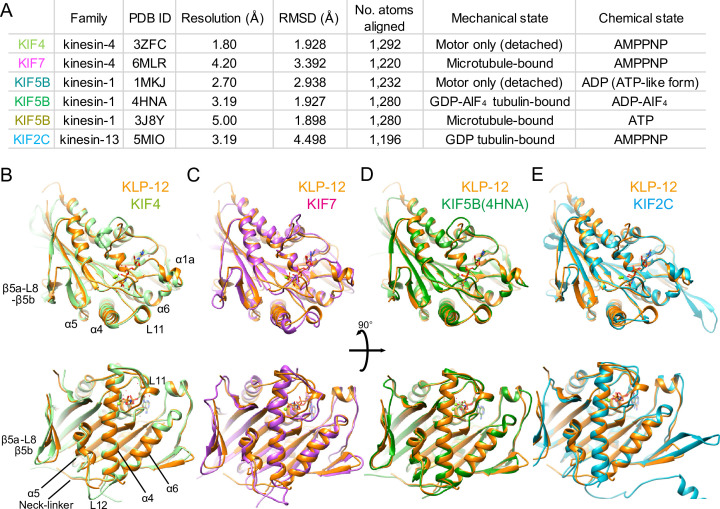
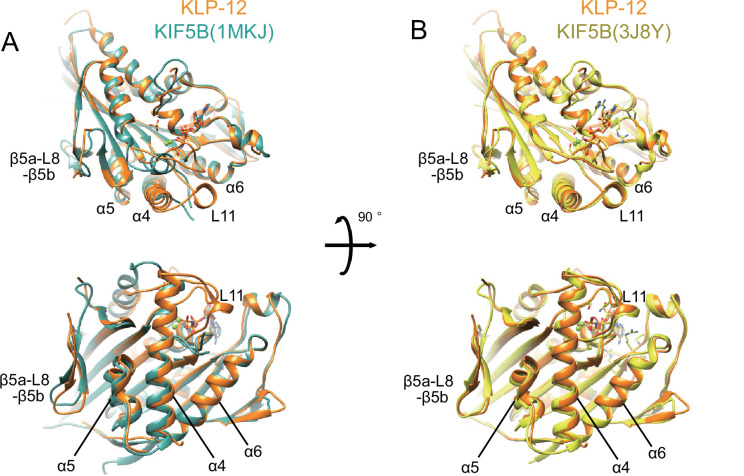
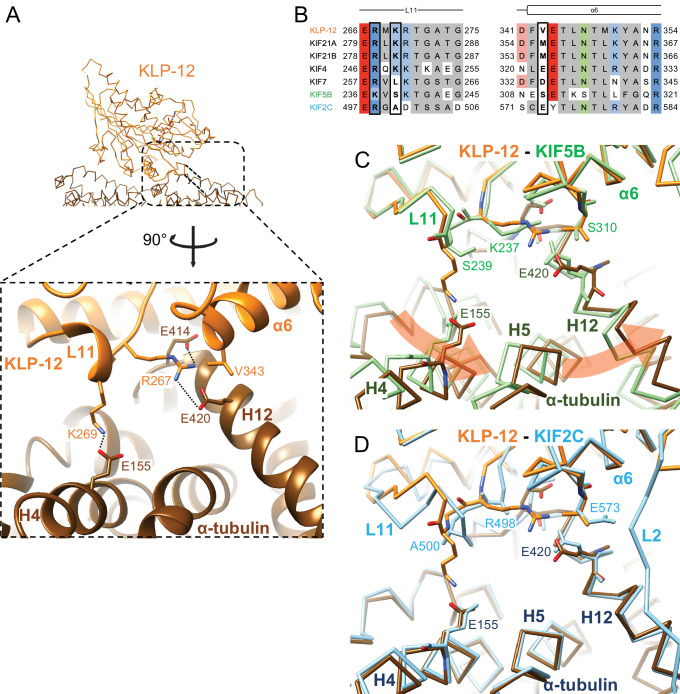
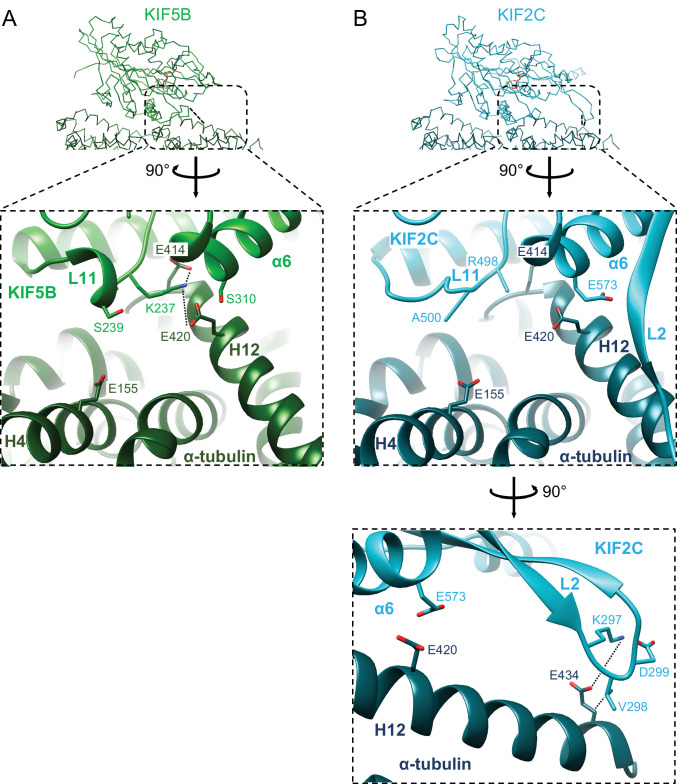
Division of Structural Medicine and Anatomy, Department of Physiology and Cell Biology, Kobe University Graduate School of Medicine, Kobe, Japan.
Division of Anesthesiology, Kobe University Graduate School of Medicine, Kobe, Japan.
Elife. 2022 Sep 6;11:e77877. doi: 10.7554/eLife.77877.
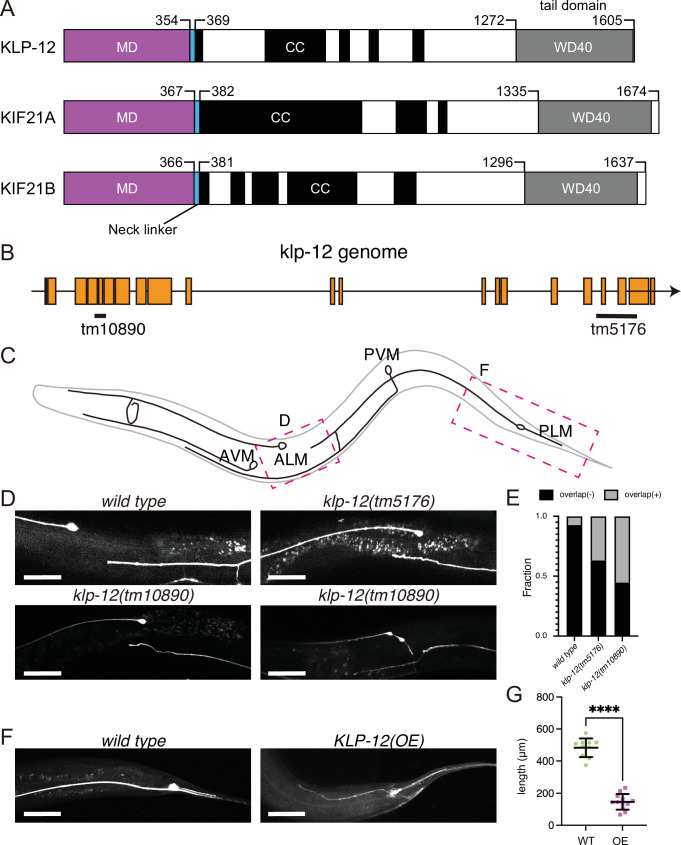
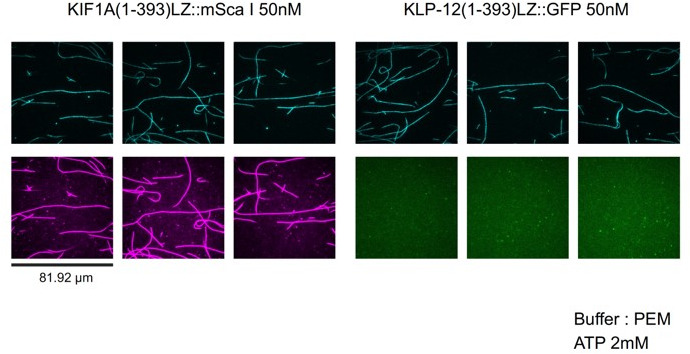
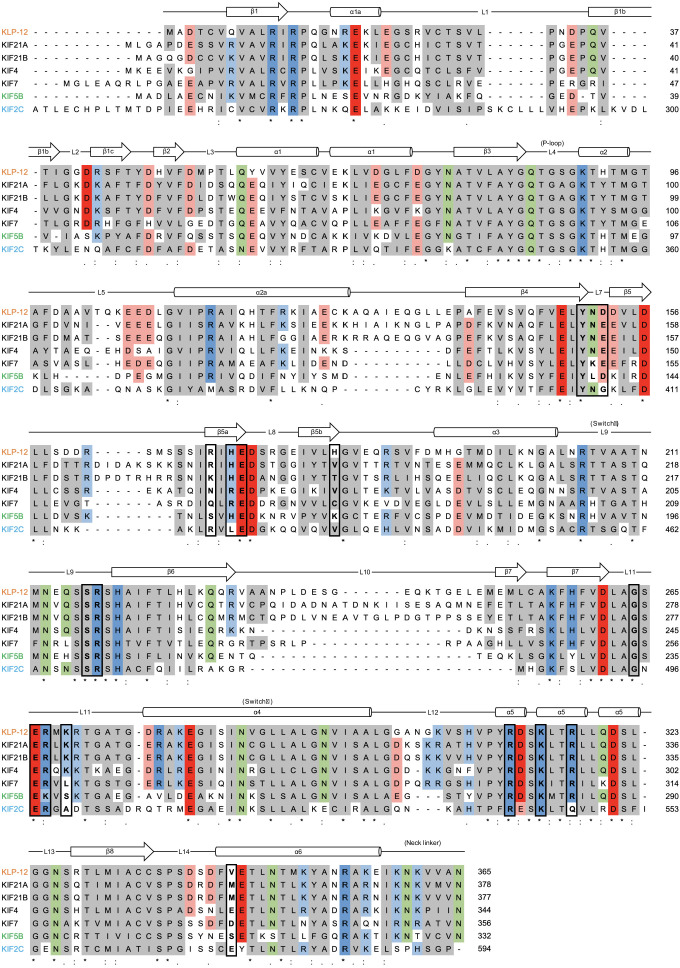
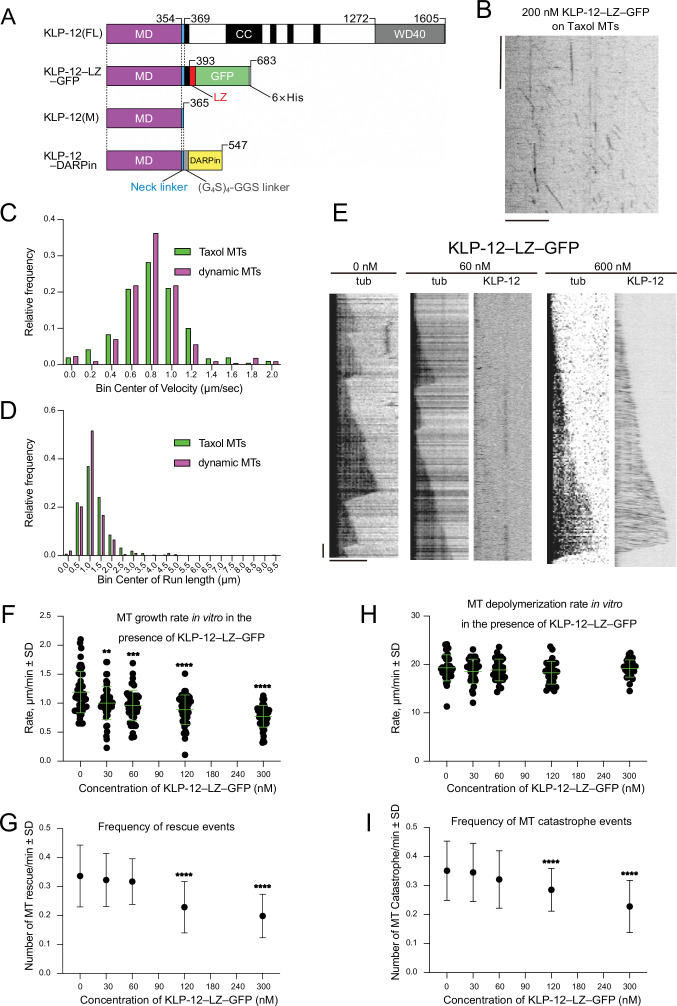
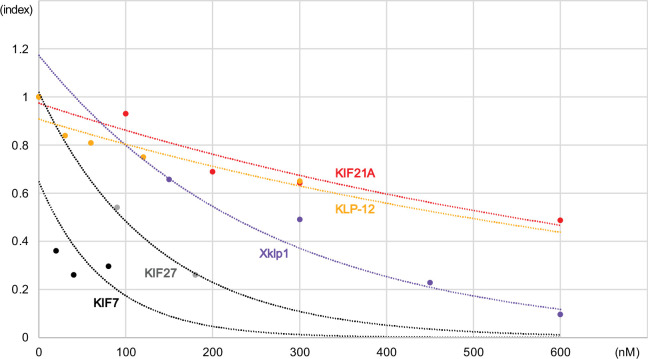
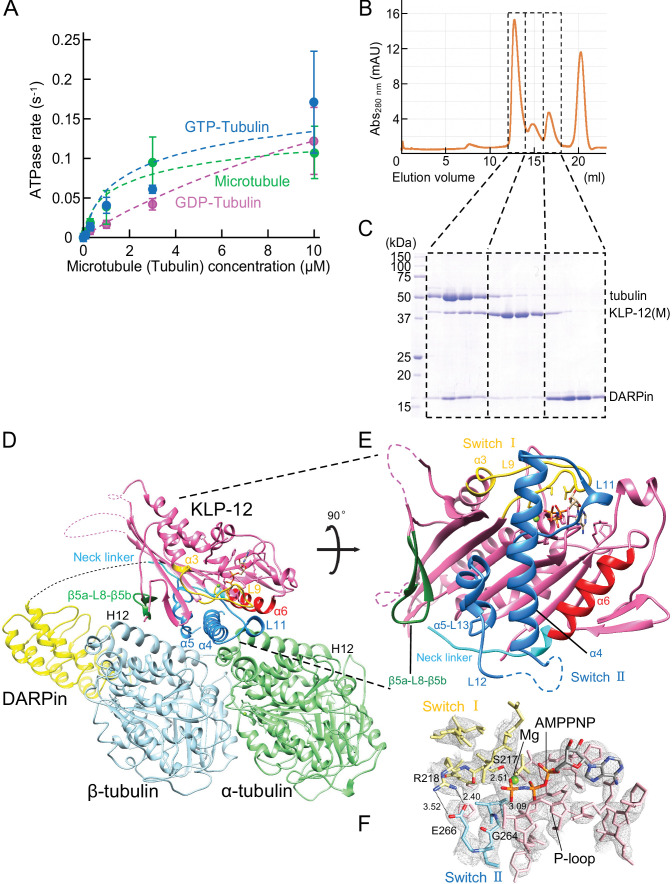
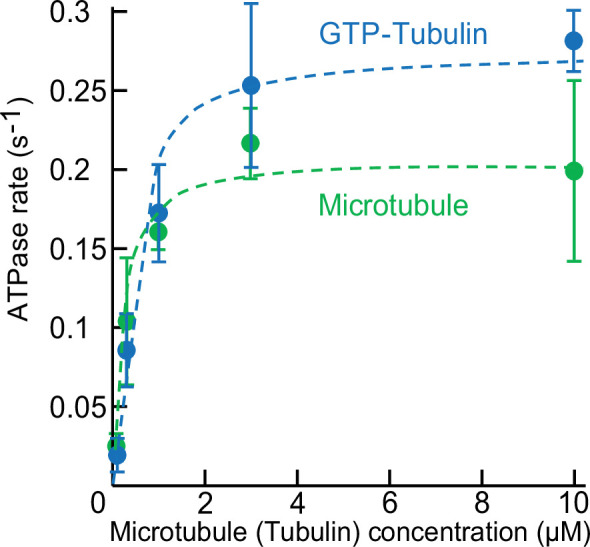
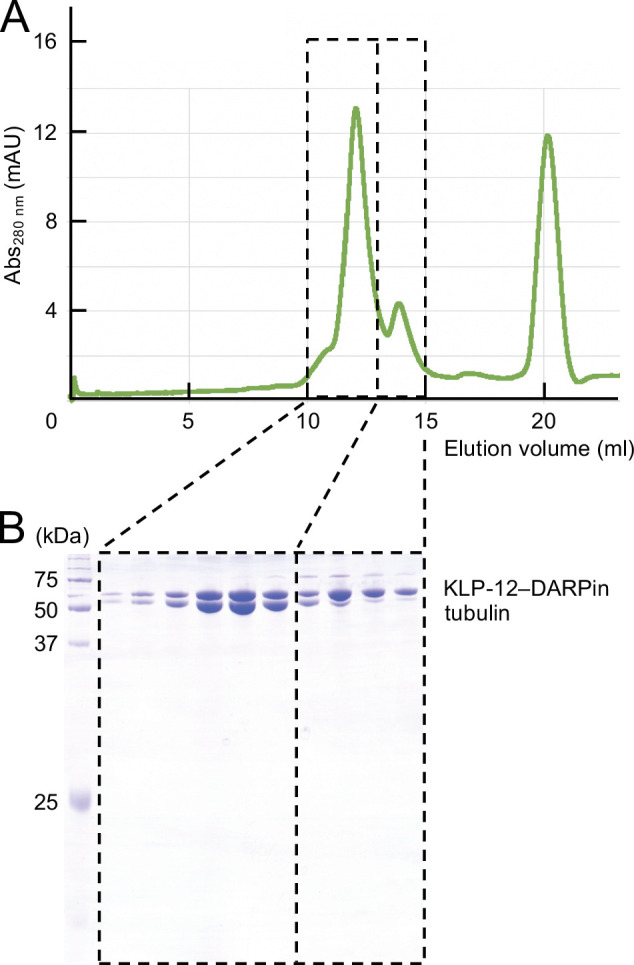
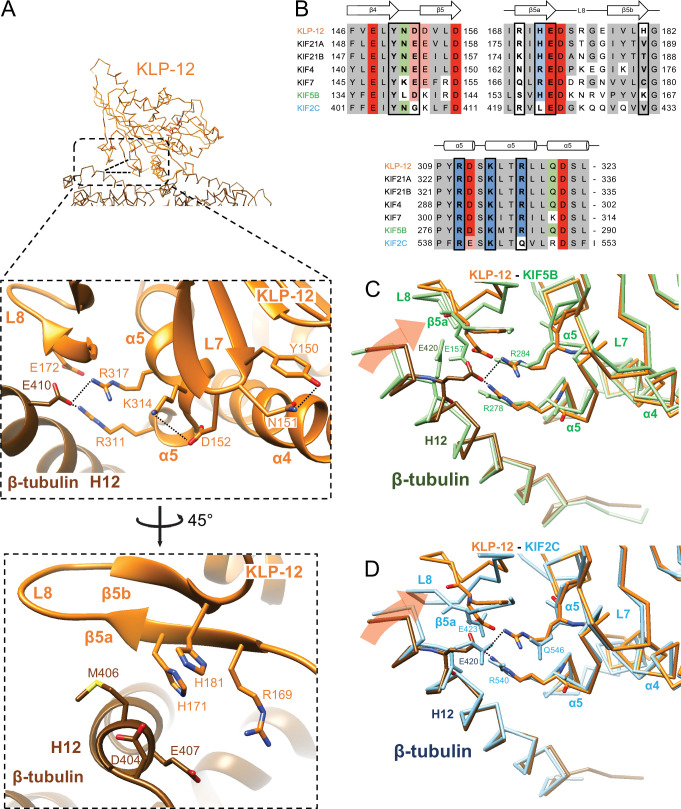
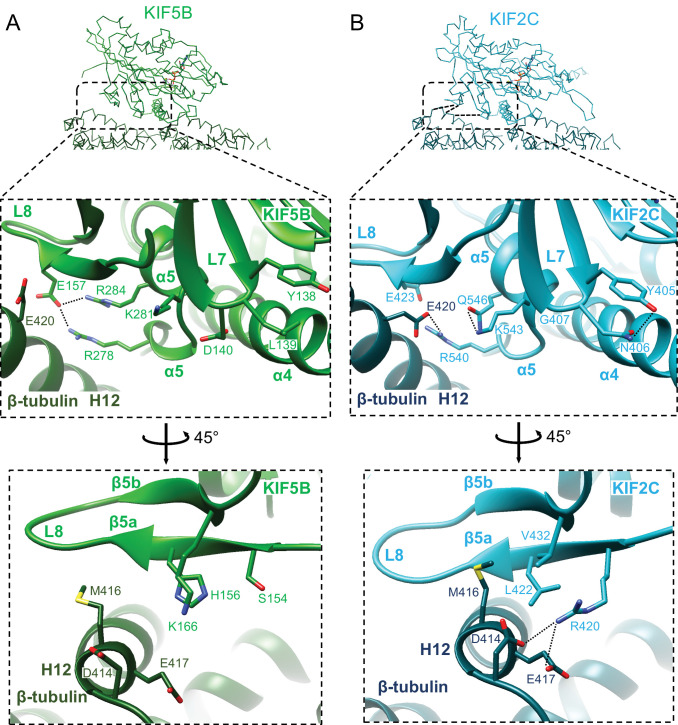
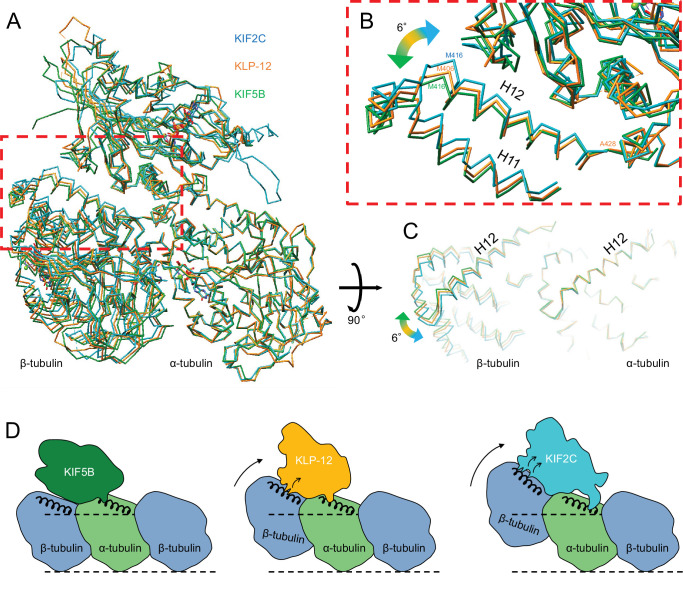
Kinesin superfamily proteins are microtubule-based molecular motors driven by the energy of ATP hydrolysis. Among them, the kinesin-4 family is a unique motor that inhibits microtubule dynamics. Although mutations of kinesin-4 cause several diseases, its molecular mechanism is unclear because of the difficulty of visualizing the high-resolution structure of kinesin-4 working at the microtubule plus-end. Here, we report that KLP-12, a kinesin-4 ortholog of KIF21A and KIF21B, is essential for proper length control of axons, and its motor domain represses microtubule polymerization in vitro. The crystal structure of the KLP-12 motor domain complexed with tubulin, which represents the high-resolution structural snapshot of the inhibition state of microtubule-end dynamics, revealed the bending effect of KLP-12 for tubulin. Comparison with the KIF5B-tubulin and KIF2C-tubulin complexes, which represent the elongation and shrinking forms of microtubule ends, respectively, showed the curvature of tubulin introduced by KLP-12 is in between them. Taken together, KLP-12 controls the proper length of axons by modulating the curvature of the microtubule ends to inhibit the microtubule dynamics.
驱动蛋白超家族蛋白是基于微管的分子马达,由 ATP 水解的能量驱动。其中,驱动蛋白-4 家族是一种独特的马达,能抑制微管动力学。尽管驱动蛋白-4 的突变会导致几种疾病,但由于难以可视化工作在微管正端的驱动蛋白-4 的高分辨率结构,其分子机制尚不清楚。在这里,我们报告说,KIF21A 和 KIF21B 的同源物 KLP-12 对于轴突的适当长度控制是必不可少的,并且它的马达结构域在体外抑制微管聚合。KLP-12 马达结构域与微管形成复合物的晶体结构,代表了微管末端动力学抑制状态的高分辨率结构快照,揭示了 KLP-12 对微管的弯曲效应。与 KIF5B-微管和 KIF2C-微管复合物的比较,分别代表微管末端的伸长和收缩形式,表明 KLP-12 引入的微管曲率介于两者之间。总之,KLP-12 通过调节微管末端的曲率来控制轴突的适当长度,从而抑制微管动力学。