Department of Biomedical and Molecular Sciences, Queen's University, Kingston, ON, K7L 3N6, Canada.
Department of Biochemistry, Albert Einstein College of Medicine, Bronx, NY, 10461, USA.
Nat Commun. 2022 Jul 20;13(1):4198. doi: 10.1038/s41467-022-31794-3.
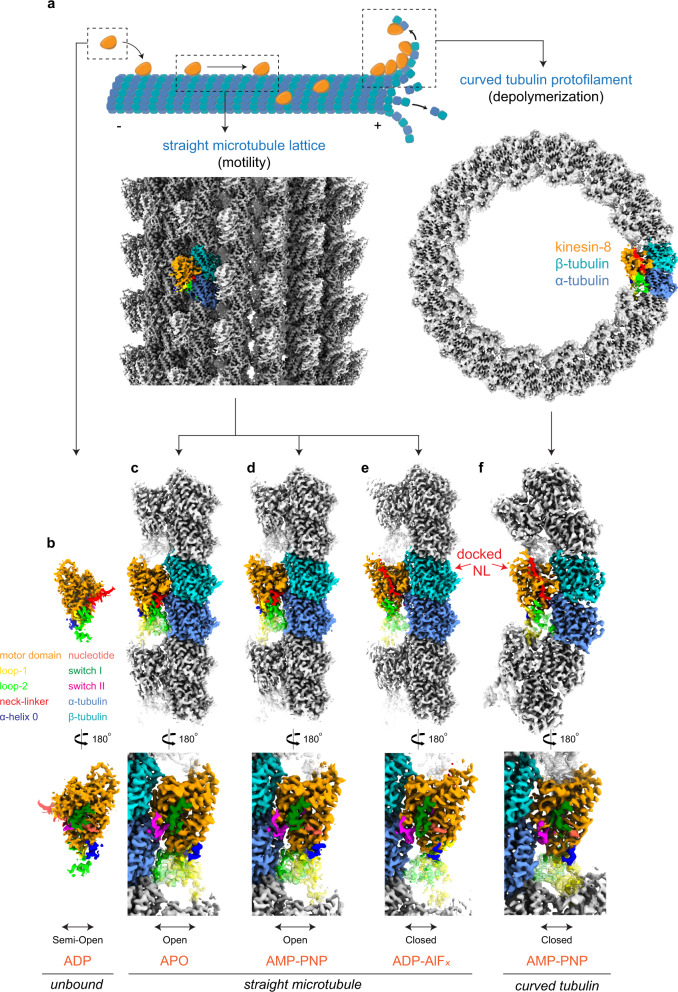
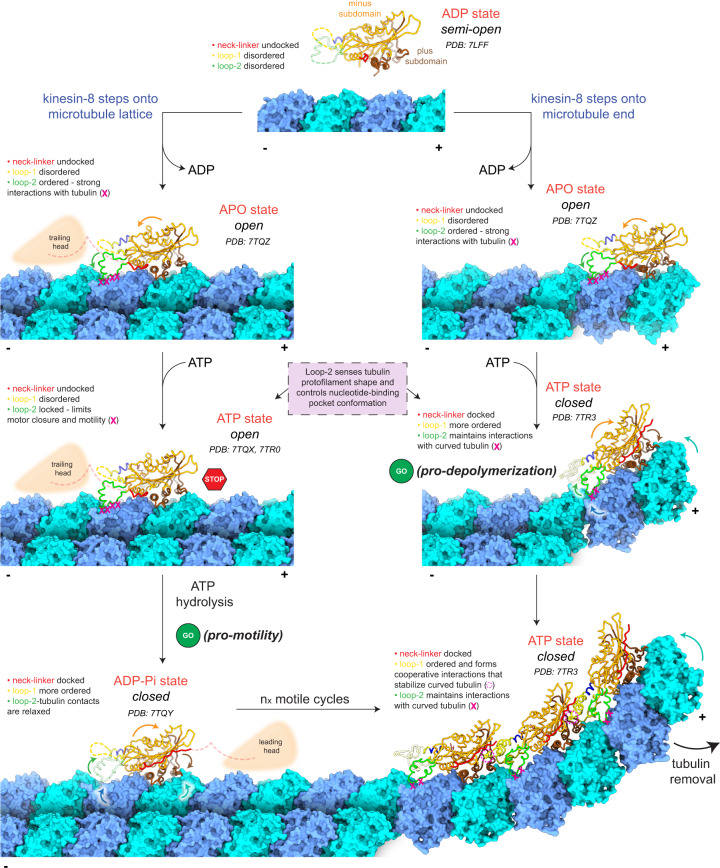
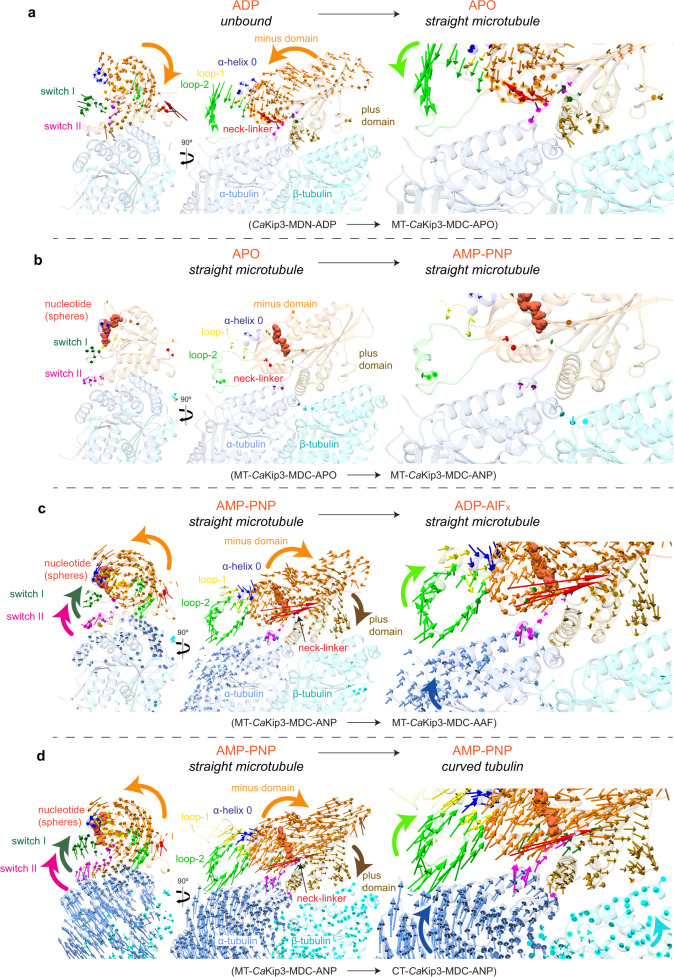
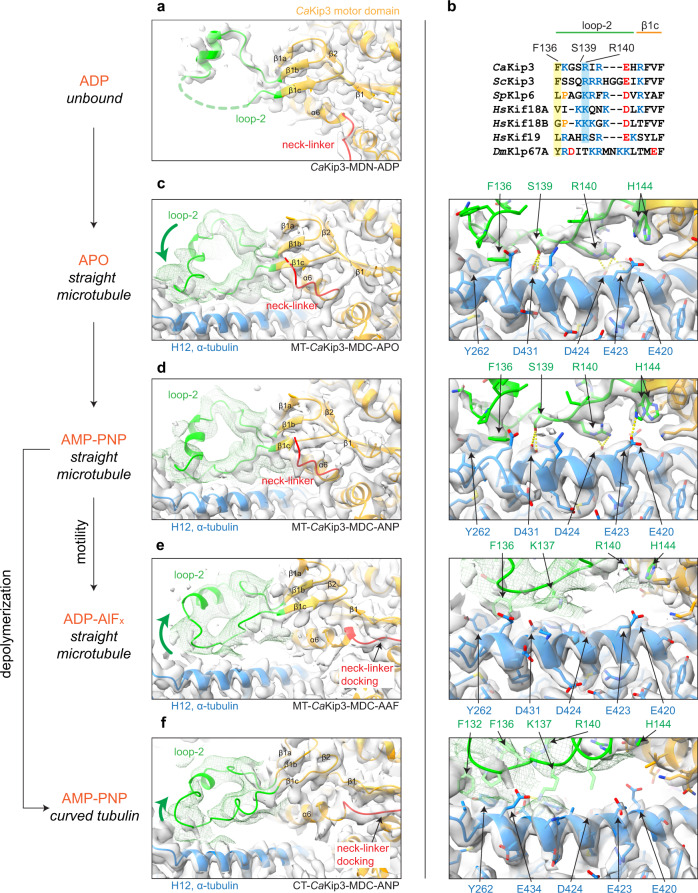
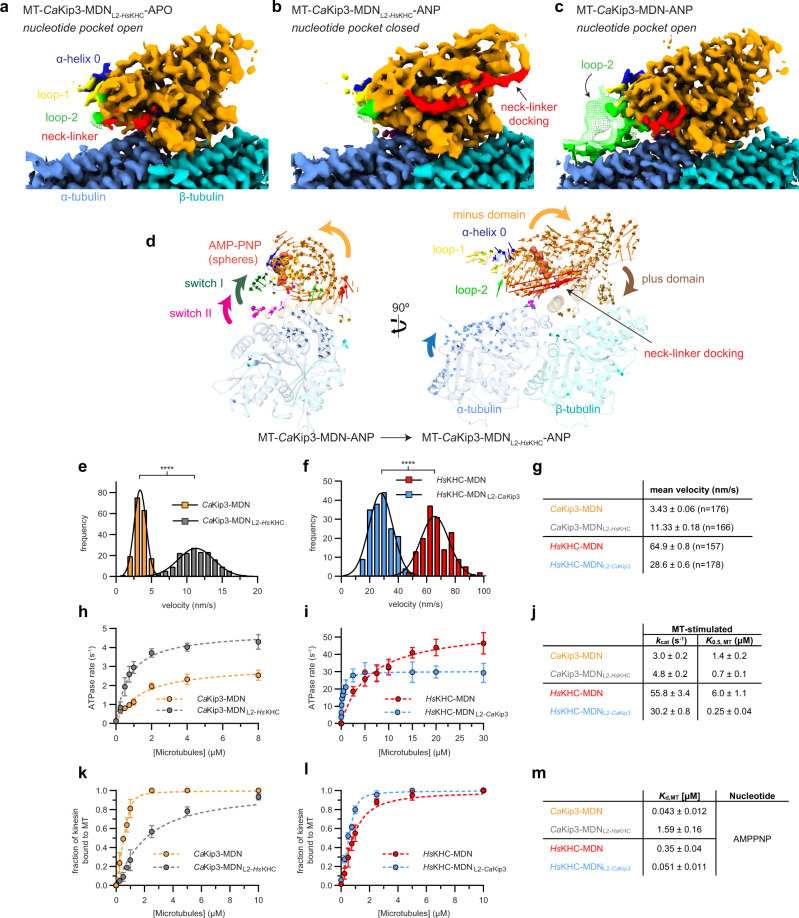
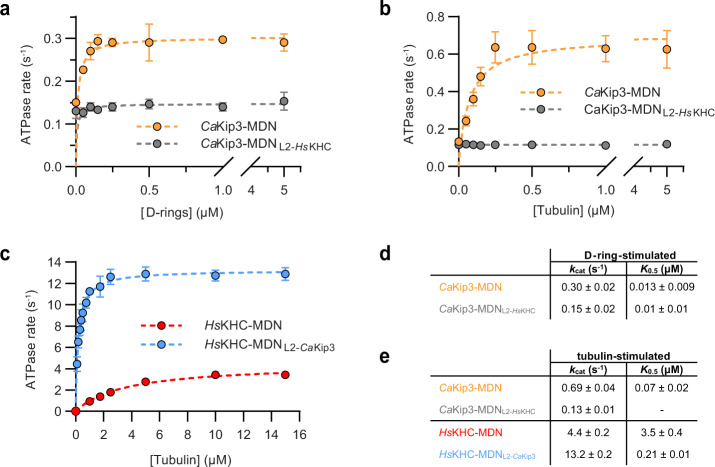
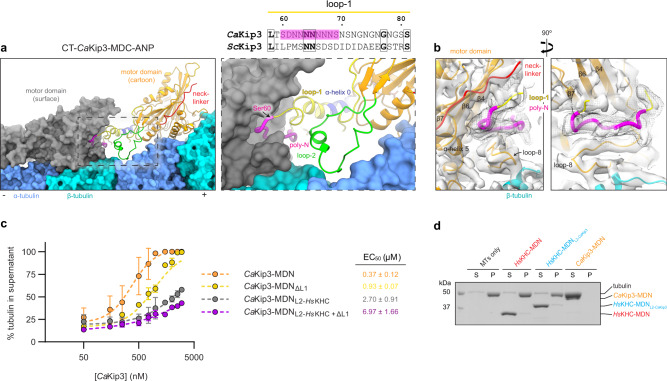
Kinesin-8s are dual-activity motor proteins that can move processively on microtubules and depolymerize microtubule plus-ends, but their mechanism of combining these distinct activities remains unclear. We addressed this by obtaining cryo-EM structures (2.6-3.9 Å) of Candida albicans Kip3 in different catalytic states on the microtubule lattice and on a curved microtubule end mimic. We also determined a crystal structure of microtubule-unbound CaKip3-ADP (2.0 Å) and analyzed the biochemical activity of CaKip3 and kinesin-1 mutants. These data reveal that the microtubule depolymerization activity of kinesin-8 originates from conformational changes of its motor core that are amplified by dynamic contacts between its extended loop-2 and tubulin. On curved microtubule ends, loop-1 inserts into preceding motor domains, forming head-to-tail arrays of kinesin-8s that complement loop-2 contacts with curved tubulin and assist depolymerization. On straight tubulin protofilaments in the microtubule lattice, loop-2-tubulin contacts inhibit conformational changes in the motor core, but in the ADP-Pi state these contacts are relaxed, allowing neck-linker docking for motility. We propose that these tubulin shape-induced alternations between pro-microtubule-depolymerization and pro-motility kinesin states, regulated by loop-2, are the key to the dual activity of kinesin-8 motors.
驱动蛋白-8s 是双活性的分子马达,它可以在微管上进行连续运动并使微管正端解聚,但它们将这两种不同活性结合起来的机制仍不清楚。我们通过获得不同催化状态下的白念珠菌 Kip3 在微管晶格上和弯曲微管末端模拟物上的冷冻电镜结构(2.6-3.9Å)来解决这个问题。我们还确定了微管非结合 CaKip3-ADP 的晶体结构(2.0Å)并分析了 CaKip3 和驱动蛋白-1 突变体的生化活性。这些数据表明,驱动蛋白-8 的微管解聚活性源于其马达核心的构象变化,该变化通过其延伸的环 2 和微管蛋白之间的动态接触而放大。在弯曲的微管末端,环 1 插入前面的马达域,形成驱动蛋白-8 的头尾排列,这些排列与弯曲的微管蛋白的环 2 接触互补,并辅助解聚。在微管晶格中的直微管原丝上,环 2-微管蛋白接触抑制马达核心的构象变化,但在 ADP-Pi 状态下,这些接触被放松,允许颈链接器对接以进行运动。我们提出,这些由微管形状诱导的在促进微管解聚和促进运动的驱动蛋白状态之间的交替,由环 2 调节,是驱动蛋白-8 马达双活性的关键。