Department of Mathematics, Columbia University, New York, United States.
Institute of Science and Technology, Maria Gugging, Austria.
Elife. 2022 Sep 26;11:e66697. doi: 10.7554/eLife.66697.
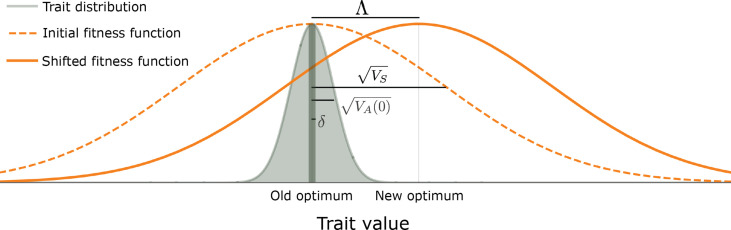
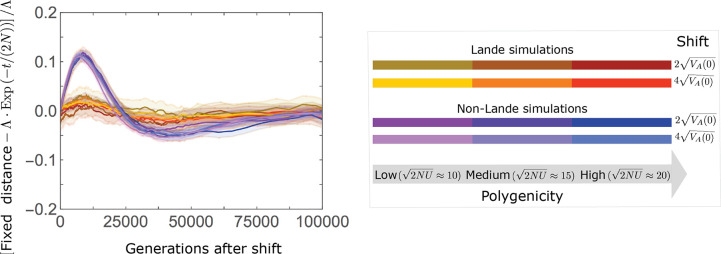
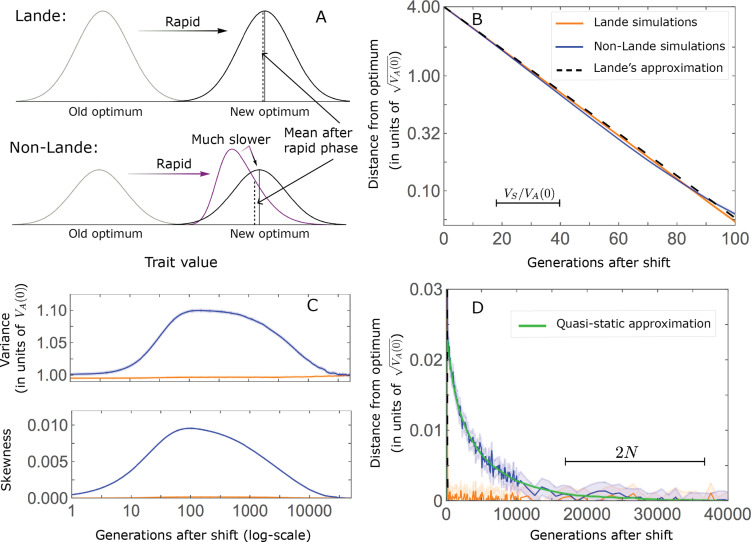
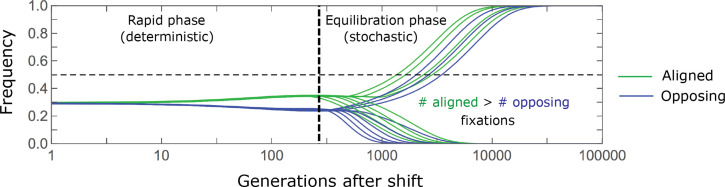
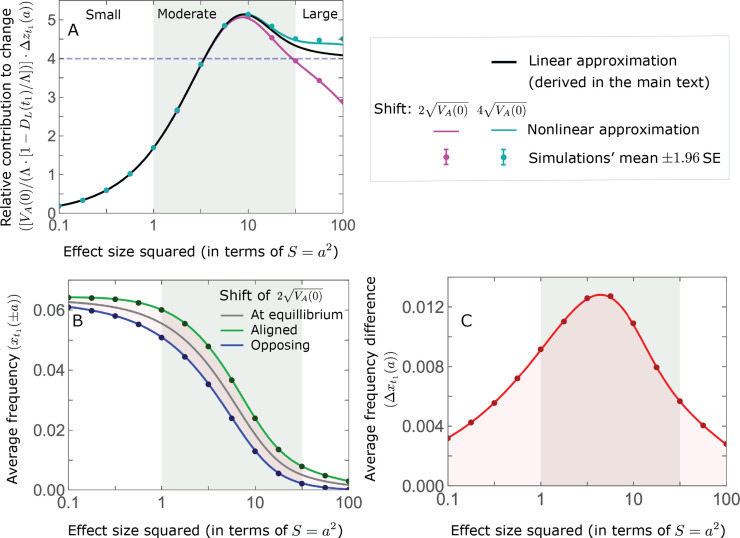
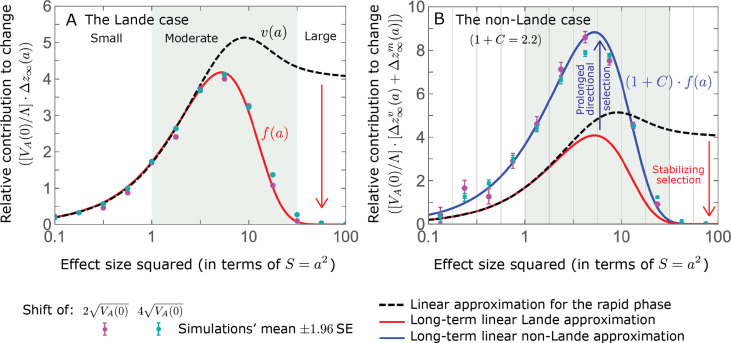
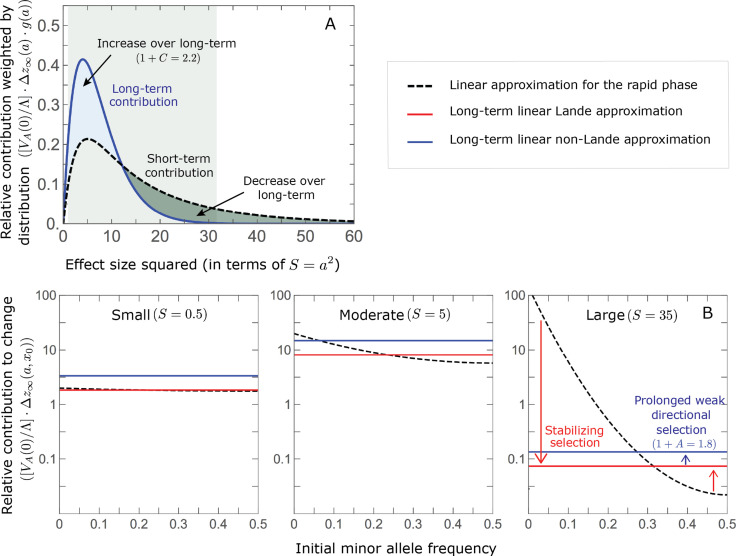
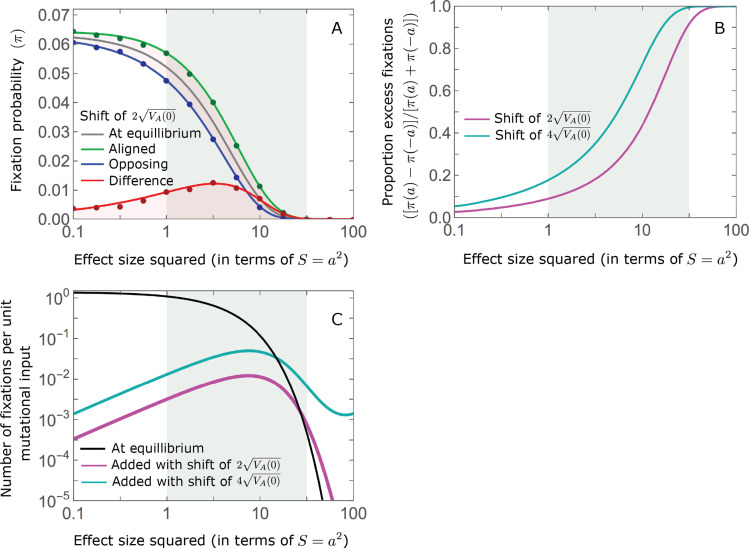
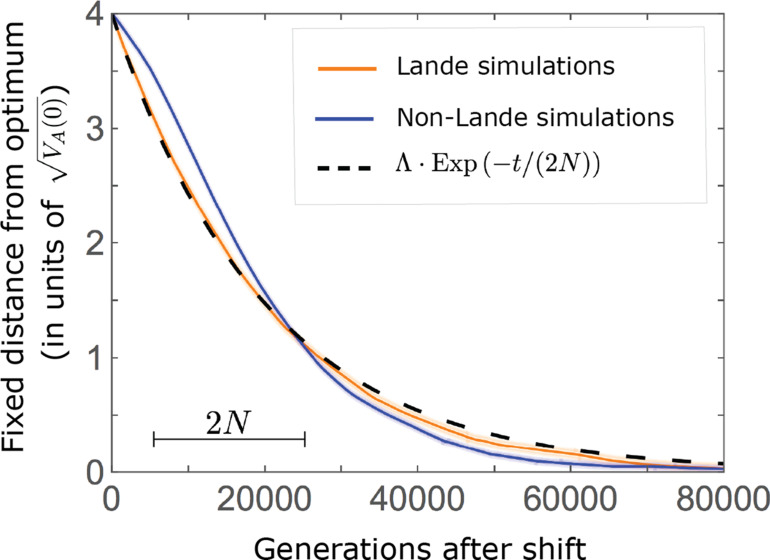
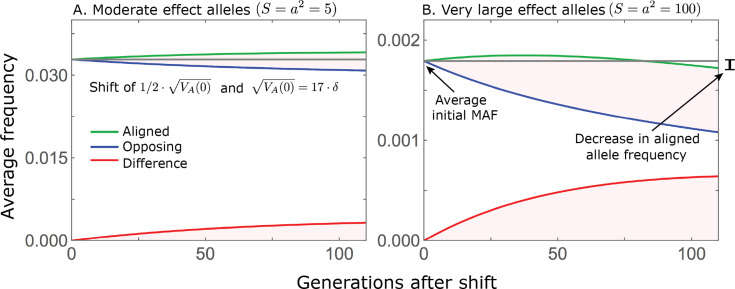
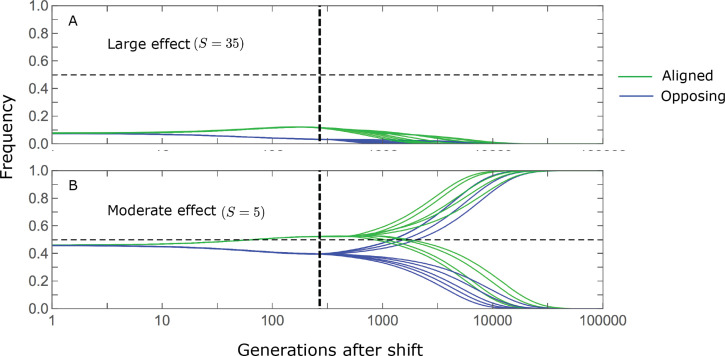
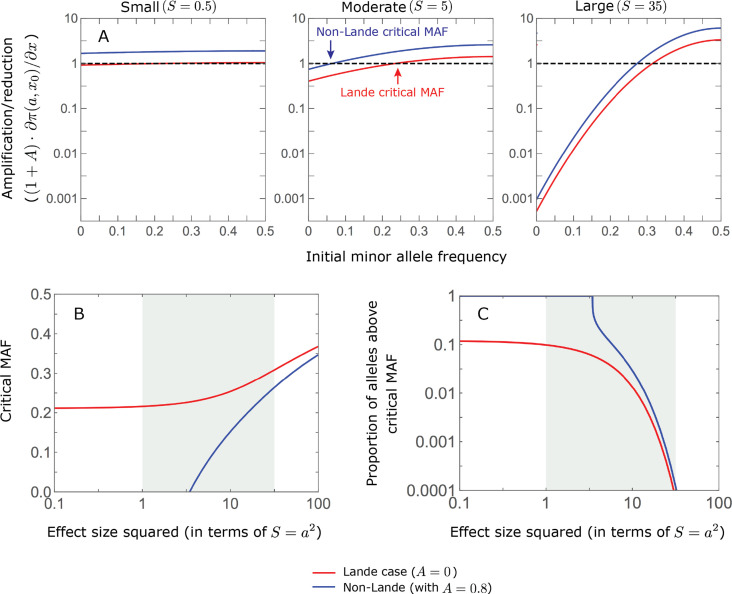
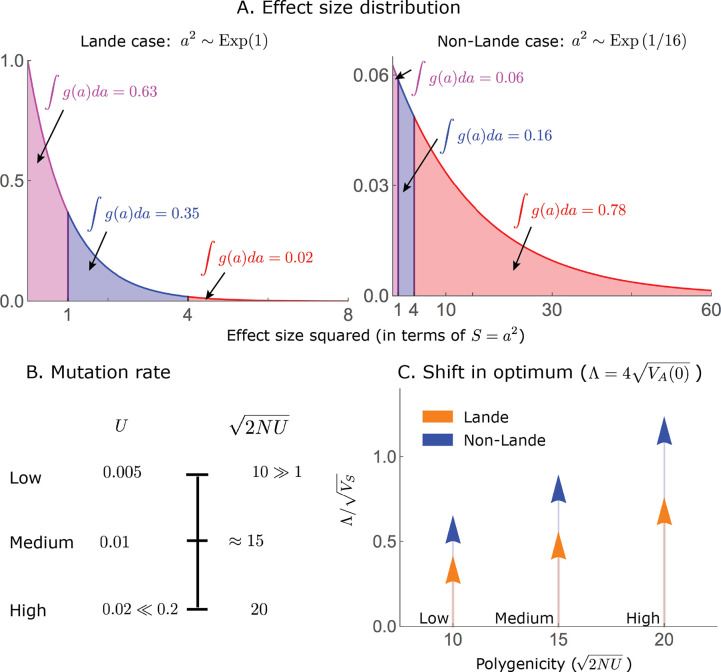
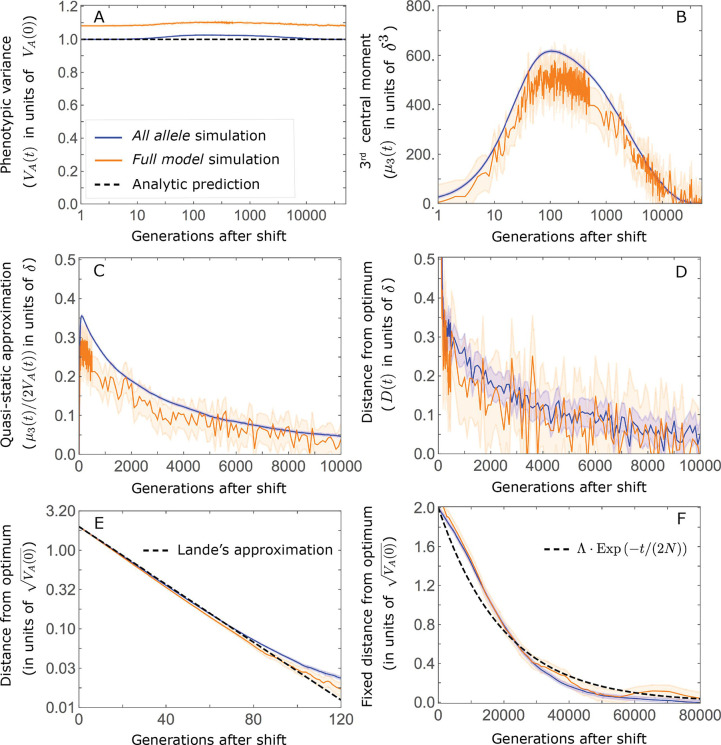
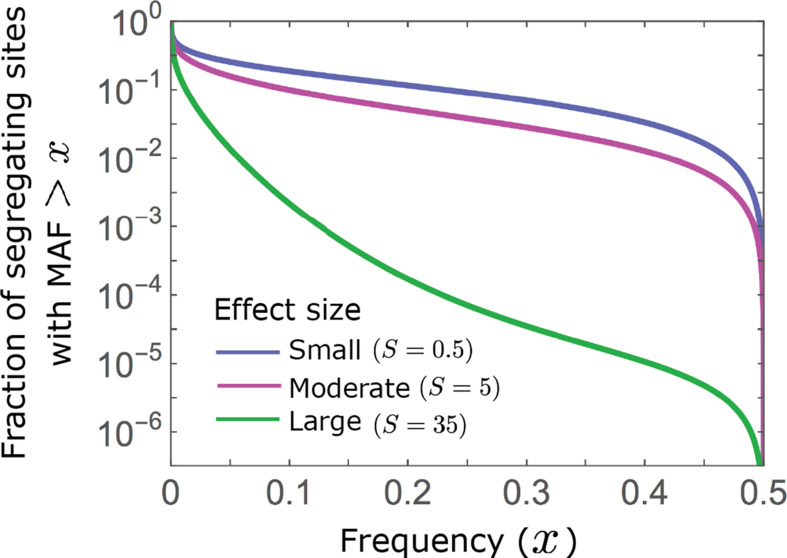
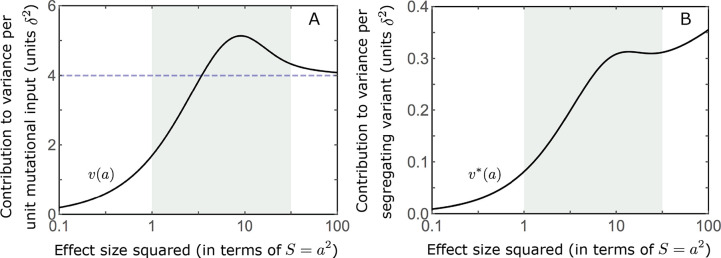
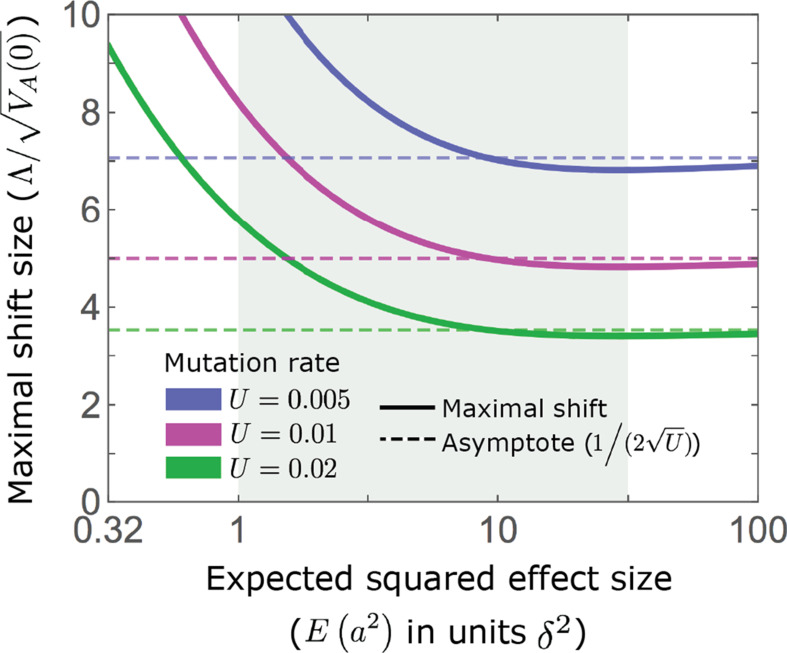
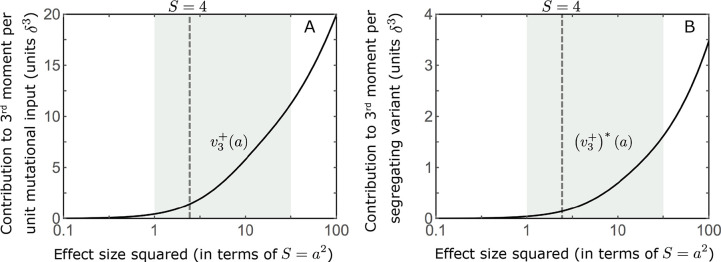
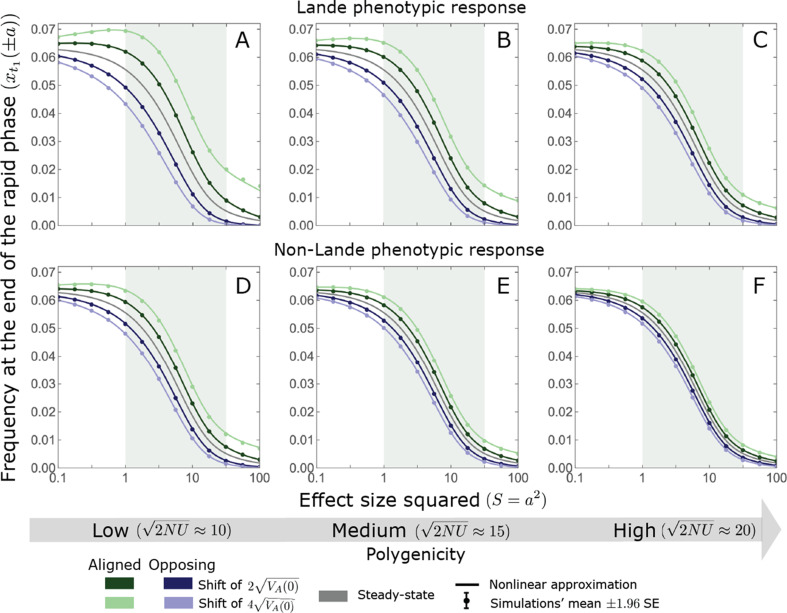
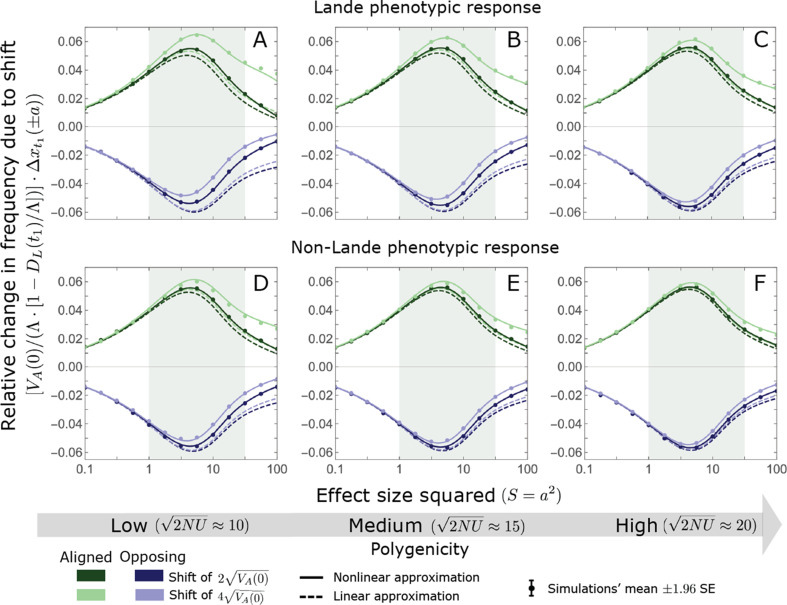
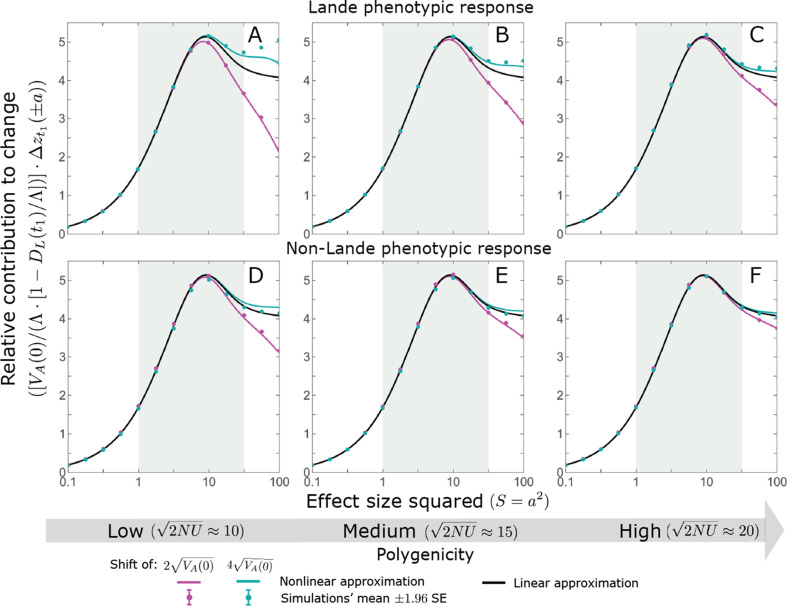
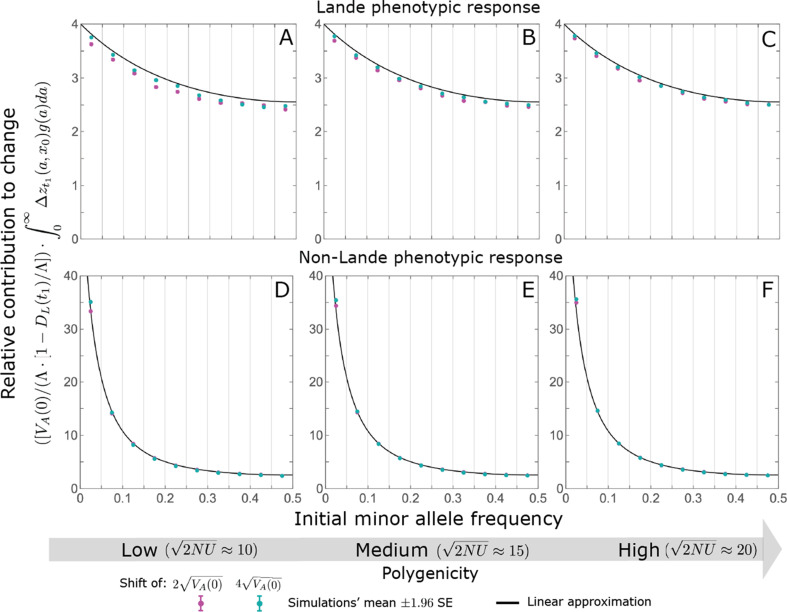
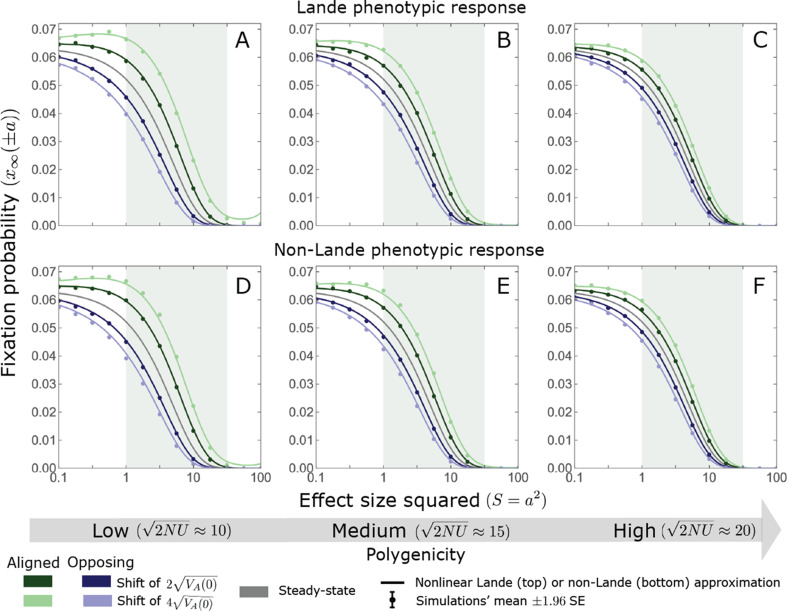
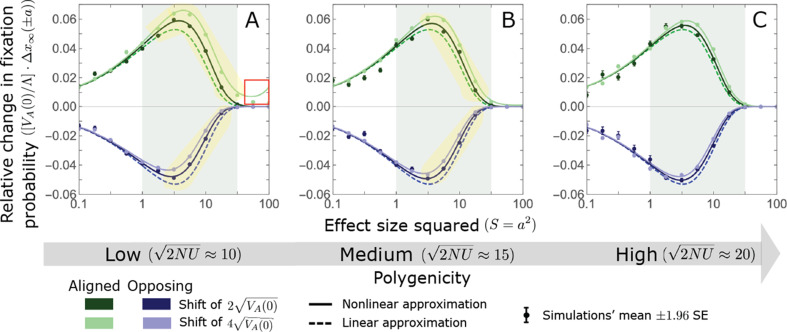
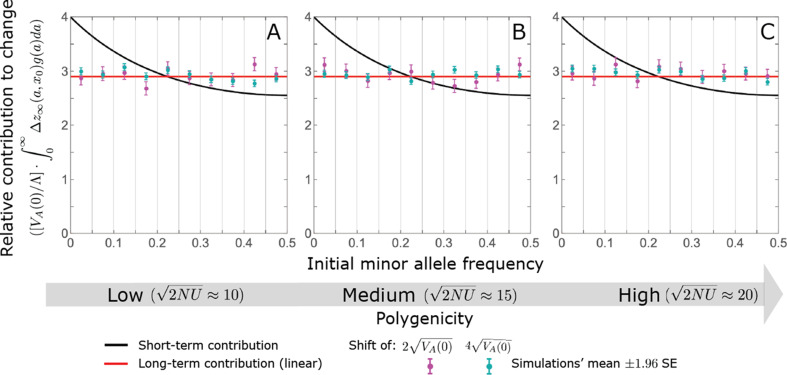
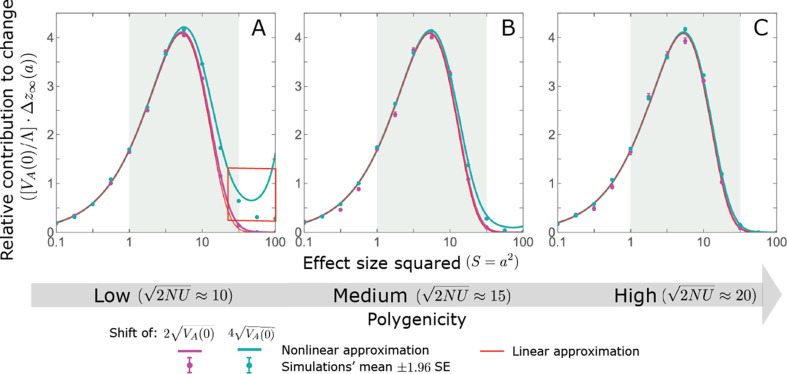
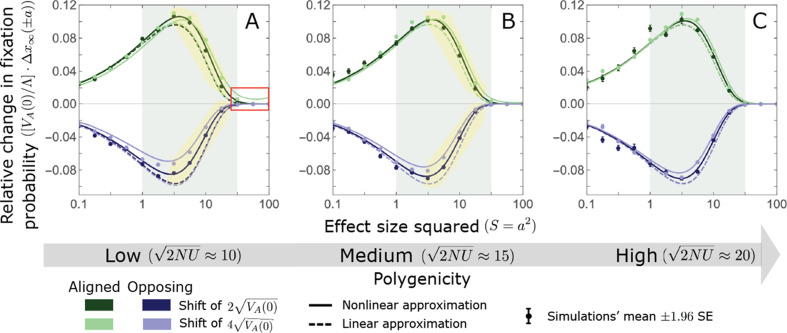
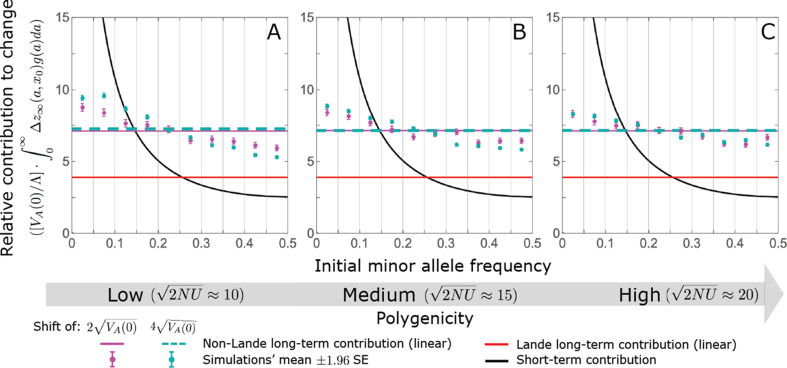
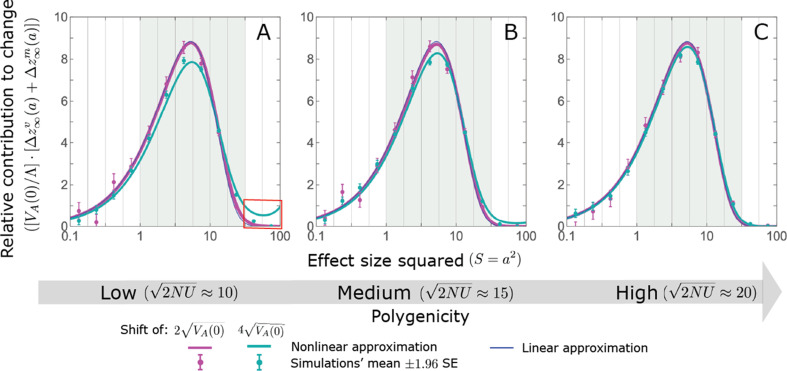
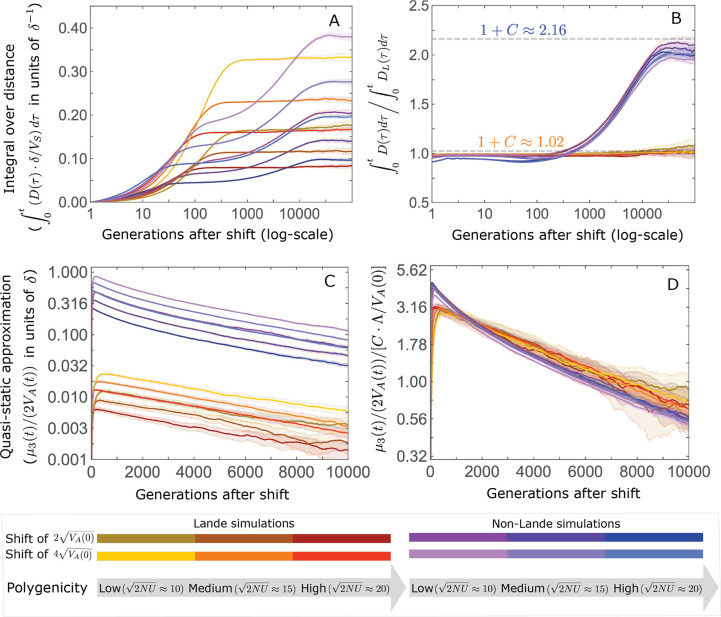
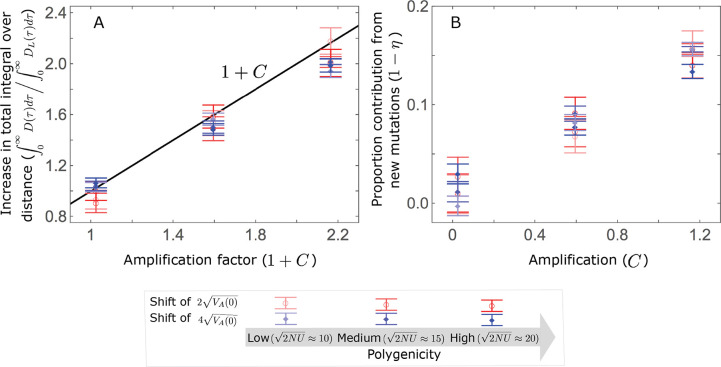
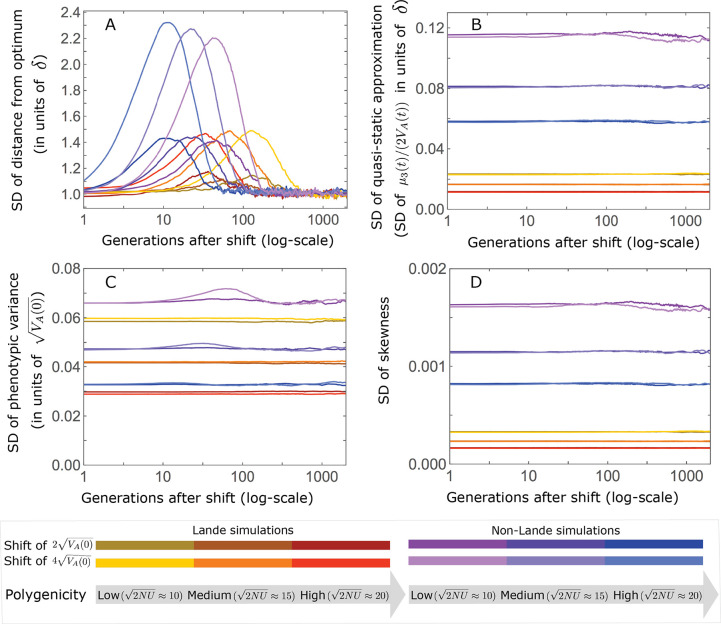
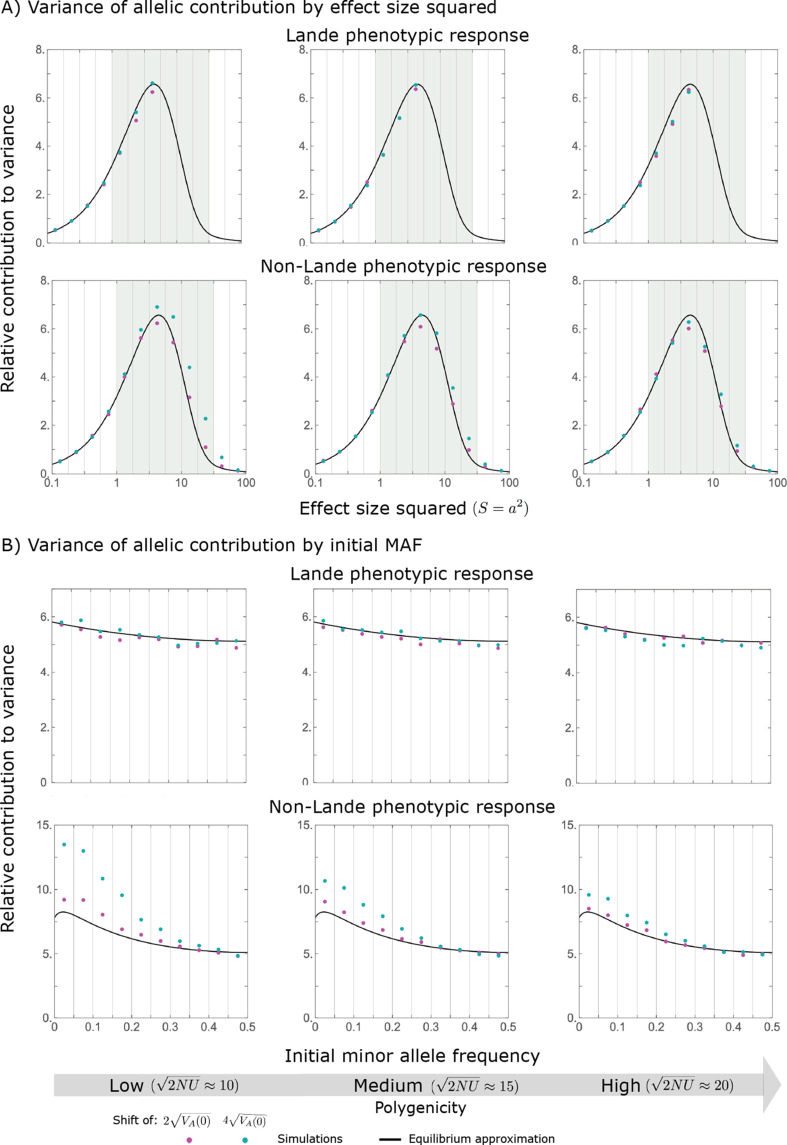
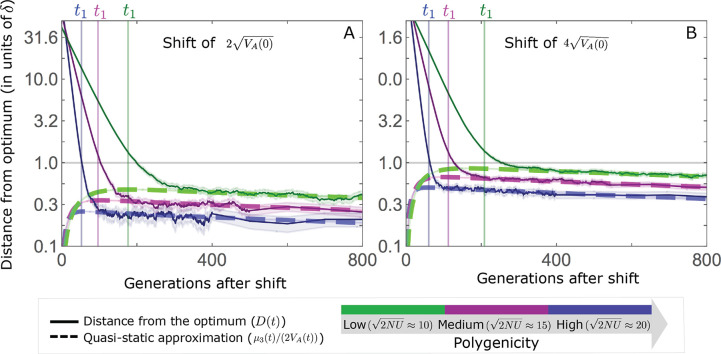
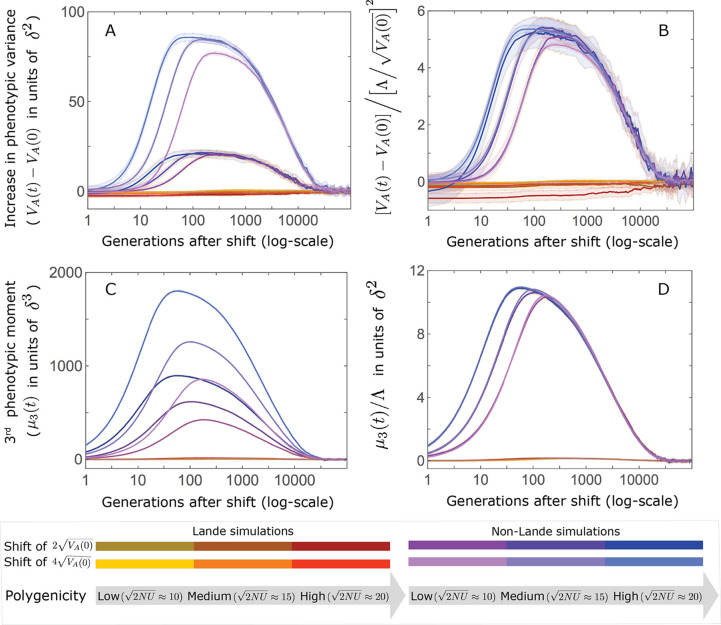
Polygenic adaptation is thought to be ubiquitous, yet remains poorly understood. Here, we model this process analytically, in the plausible setting of a highly polygenic, quantitative trait that experiences a sudden shift in the fitness optimum. We show how the mean phenotype changes over time, depending on the effect sizes of loci that contribute to variance in the trait, and characterize the allele dynamics at these loci. Notably, we describe the two phases of the allele dynamics: The first is a rapid phase, in which directional selection introduces small frequency differences between alleles whose effects are aligned with or opposed to the shift, ultimately leading to small differences in their probability of fixation during a second, longer phase, governed by stabilizing selection. As we discuss, key results should hold in more general settings and have important implications for efforts to identify the genetic basis of adaptation in humans and other species.
多基因适应被认为是普遍存在的,但仍知之甚少。在这里,我们在一个高度多基因、数量性状的合理假设环境中对这个过程进行了分析,这个性状经历了适应最优值的突然转变。我们展示了随着时间的推移,表型均值是如何变化的,这取决于对性状的变异有贡献的基因座的效应大小,并描述了这些基因座上的等位基因动态。值得注意的是,我们描述了等位基因动态的两个阶段:第一阶段是一个快速阶段,在这个阶段,定向选择在效应与转变一致或相反的等位基因之间引入了较小的频率差异,最终导致它们在第二阶段,由稳定选择控制的更长的固定概率差异较小。正如我们所讨论的,关键结果应该在更一般的环境中成立,并对人类和其他物种适应的遗传基础的识别工作有重要意义。