Kojima Kenji K, Bao Weidong
Genetic Information Research Institute, Cupertino, CA, 95014, USA.
Mob DNA. 2022 Oct 22;13(1):24. doi: 10.1186/s13100-022-00281-3.
DNA transposons are ubiquitous components of eukaryotic genomes. A major group of them encode a DDD/E transposase and contain terminal inverted repeats (TIRs) of varying lengths. The Kolobok superfamily of DNA transposons has been found in a wide spectrum of organisms.
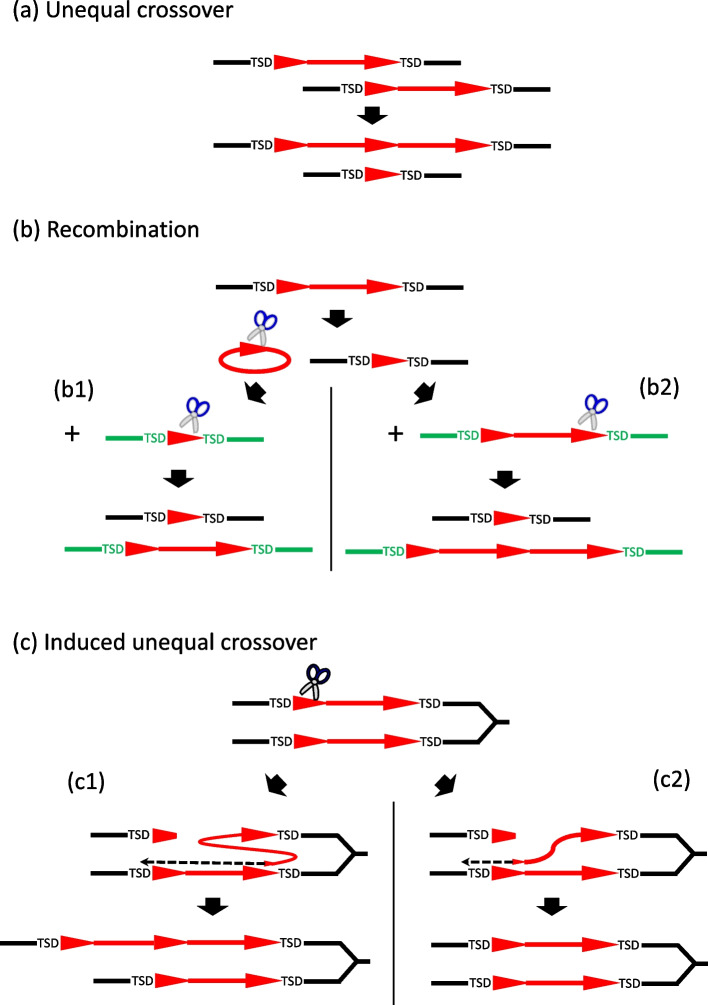
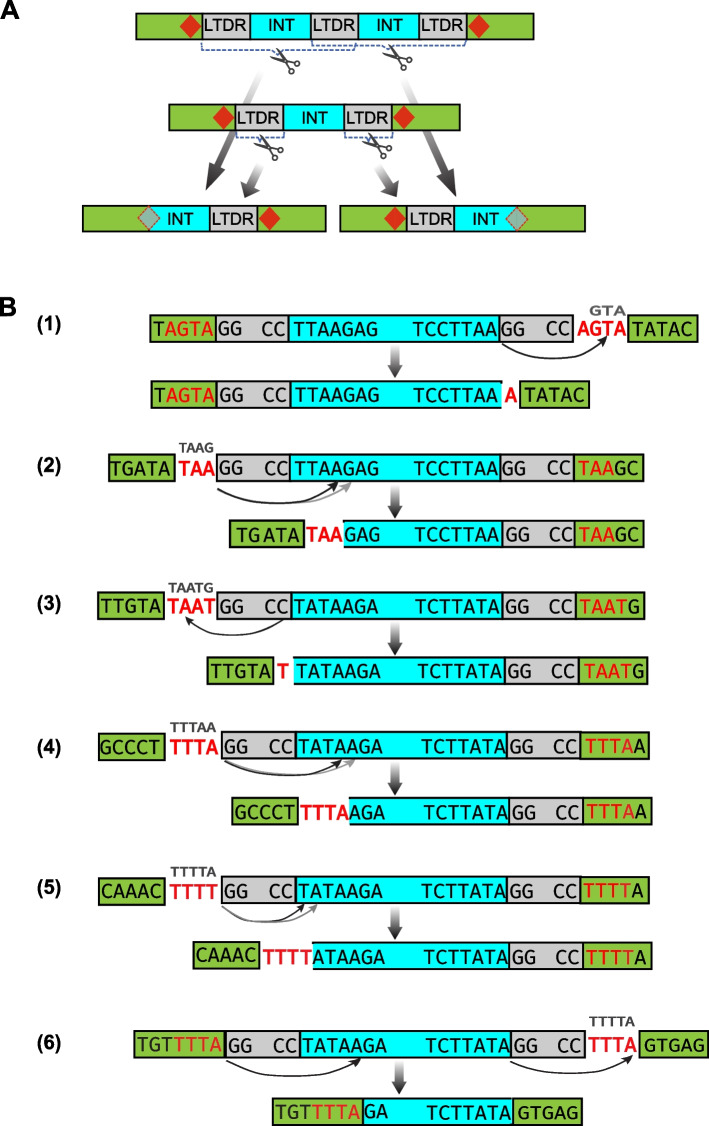
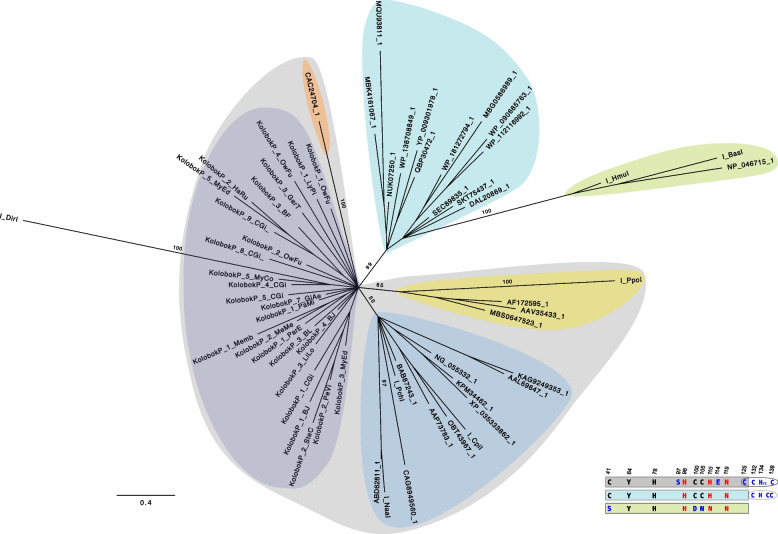
Here we report a new Kolobok lineage, designated KolobokP. They were identified in 7 animal phyla (Mollusca, Phoronida, Annelida, Nemertea, Bryozoa, Chordata, and Echinodermata), and are especially rich in bivalves. Unlike other Kolobok families, KolobokP adopts a composite-like architecture: an internal region (INT) flanked by two long terminal direct repeats (LTDRs), which exhibit their own short terminal inverted repeats ranging up to 18 bps. The excision of LTDRs was strongly suggested. The LTDR lengths seem to be constrained to be either around 450-bp or around 660-bp. The internal region encodes a DDD/E transposase and a small His-Me finger nuclease, which likely originated from the homing endonuclease encoded by a group I intron from a eukaryotic species. The architecture of KolobokP resembles composite DNA transposons, usually observed in bacterial genomes, and long terminal repeat (LTR) retrotransposons. In addition to this monomeric LTDR-INT-LTDR structure, plenty of solo LTDRs and multimers represented as (LTDR-INT)-LTDR are also observed. Our structural and phylogenetic analysis supported the birth of KolobokP in the late stage of the Kolobok evolution. We propose KolobokP families propagate themselves in two ways: the canonical transposition catalyzed by their transposase and the sequence-specific cleavage by their endonuclease followed by the multimerization through the unequal crossover.
The presence of homing endonuclease and long terminal direct repeats of KolobokP families suggest their unique dual replication mechanisms: transposition and induced unequal crossover.
DNA转座子是真核生物基因组中普遍存在的组成部分。其中一大类编码DDD/E转座酶,并含有不同长度的末端反向重复序列(TIRs)。DNA转座子的Kolobok超家族已在广泛的生物体中被发现。
在此我们报告一个新的Kolobok谱系,命名为KolobokP。它们在7个动物门(软体动物门、帚虫动物门、环节动物门、纽形动物门、苔藓虫纲、脊索动物门和棘皮动物门)中被鉴定出来,在双壳贝类中尤为丰富。与其他Kolobok家族不同,KolobokP采用类似复合结构的架构:一个内部区域(INT)两侧是两个长末端正向重复序列(LTDRs),这些LTDRs有自身短至18个碱基对的末端反向重复序列。强烈提示LTDRs会发生切除。LTDR的长度似乎被限制在约450碱基对或约660碱基对左右。内部区域编码一个DDD/E转座酶和一个小的His-Me指状核酸酶,后者可能起源于真核生物物种I组内含子编码的归巢内切核酸酶。KolobokP的架构类似于通常在细菌基因组中观察到的复合DNA转座子以及长末端重复(LTR)逆转录转座子。除了这种单体LTDR-INT-LTDR结构外,还观察到大量单独的LTDRs以及以(LTDR-INT)-LTDR形式存在的多聚体。我们的结构和系统发育分析支持KolobokP在Kolobok进化后期诞生。我们提出KolobokP家族以两种方式自我繁殖:由其转座酶催化的典型转座以及由其内切核酸酶进行的序列特异性切割,随后通过不等交换进行多聚化。
KolobokP家族中归巢内切核酸酶和长末端正向重复序列的存在表明它们具有独特的双重复制机制:转座和诱导不等交换。