Martin Eliza C, Vicari Célia, Tsakou-Ngouafo Louis, Pontarotti Pierre, Petrescu Andrei J, Schatz David G
1Department of Bioinformatics and Structural Biochemistry, Institute of Biochemistry of the Romanian Academy, Splaiul Independentei 296, 060031 Bucharest, Romania.
2Evolutionary biology team, Aix Marseille Université IRD, APHM, MEPHI, IHU Méditerranée Infection, Marseille, France.
Mob DNA. 2020 May 6;11:17. doi: 10.1186/s13100-020-00214-y. eCollection 2020.
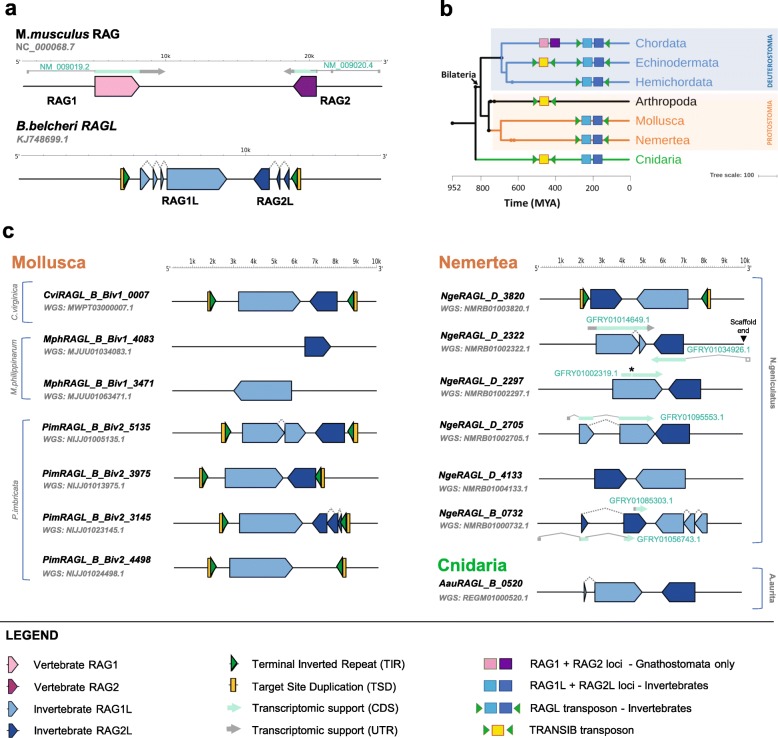
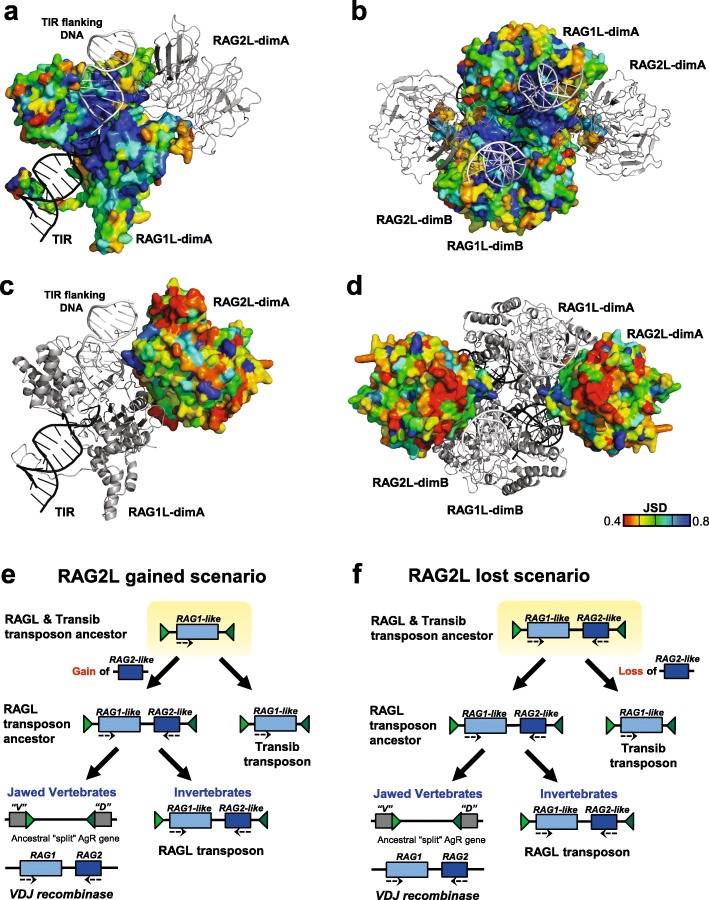
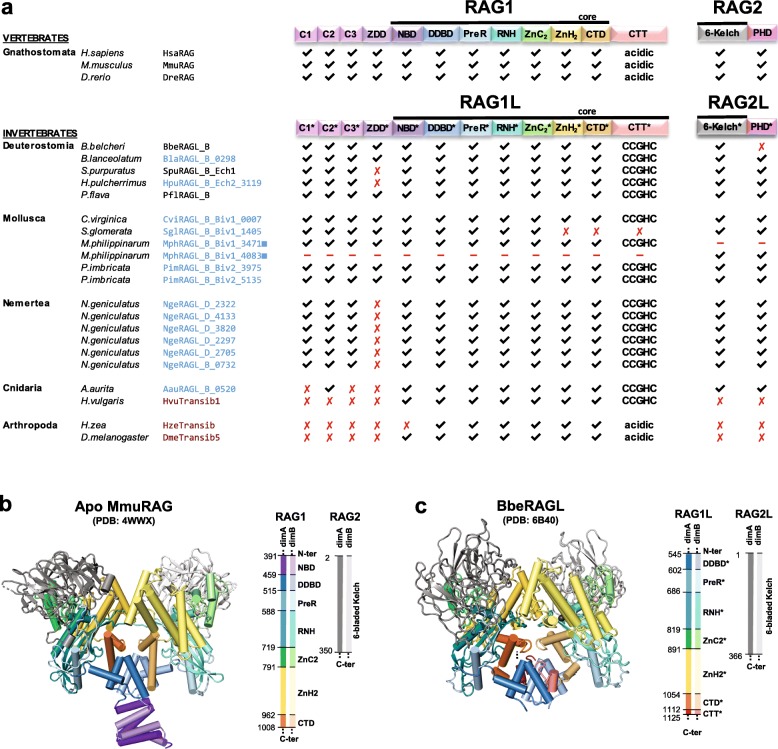
V(D) J recombination is essential for adaptive immunity in jawed vertebrates and is initiated by the RAG1-RAG2 endonuclease. The and genes are thought to have evolved from a (RAG-like) transposon containing convergently-oriented () and genes. Elements resembling this presumptive evolutionary precursor have thus far only been detected convincingly in deuterostomes, leading to the model that the transposon first appeared in an early deuterostome.
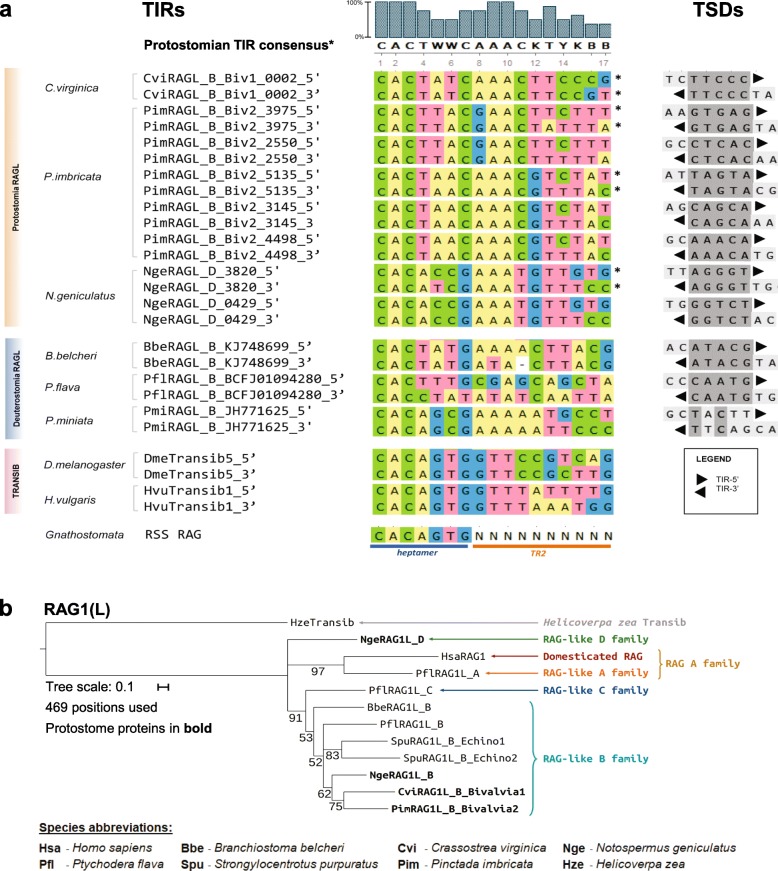
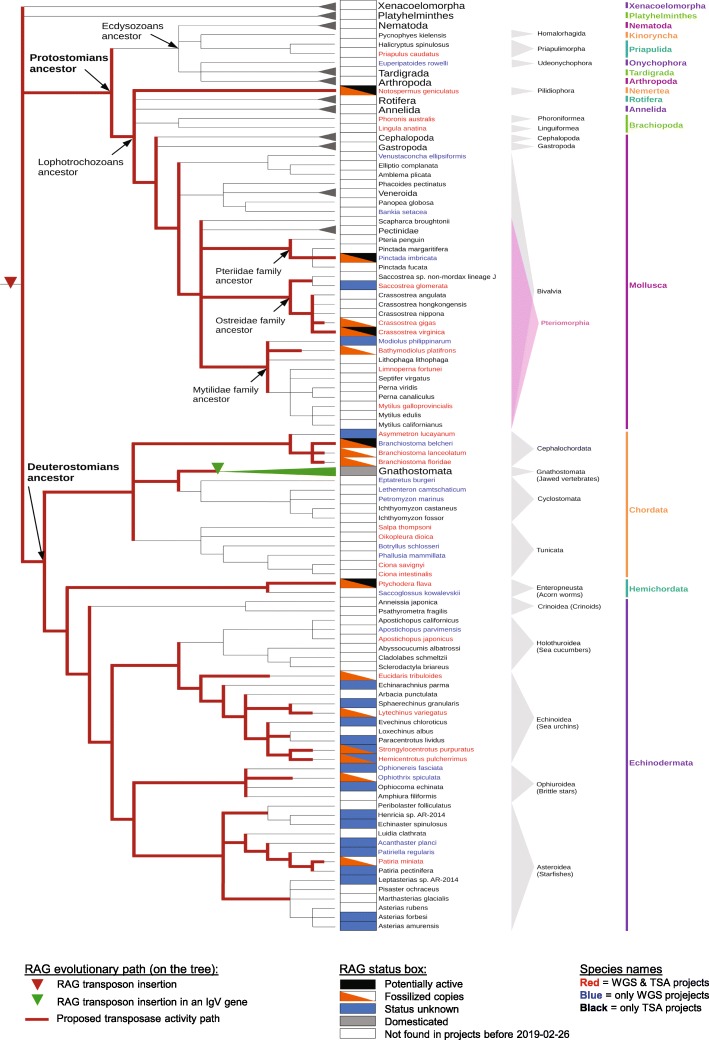
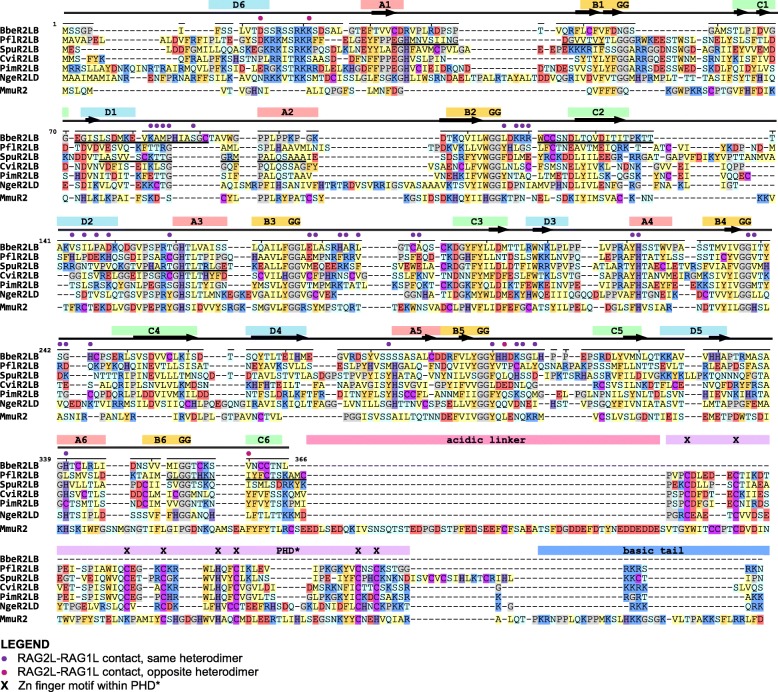
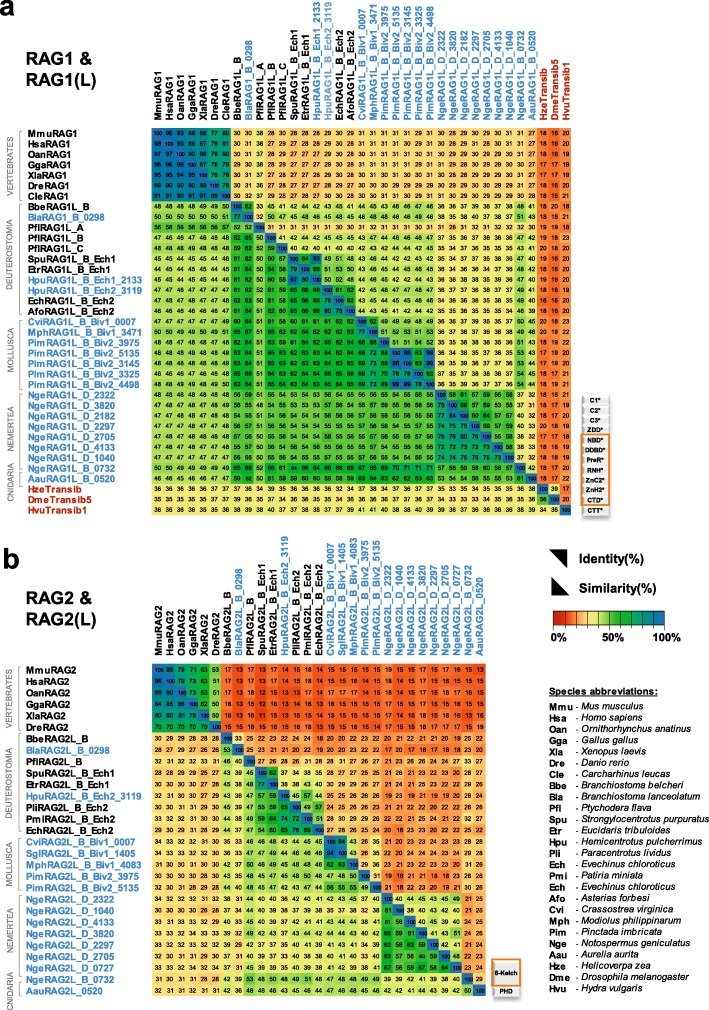
We have identified numerous transposons in the genomes of protostomes, including oysters and mussels (phylum Mollusca) and a ribbon worm (phylum Nemertea), and in the genomes of several cnidarians. Phylogenetic analyses are consistent with vertical evolution of transposons within the Bilateria clade and with its presence in the bilaterian ancestor. Many of the transposons identified in protostomes are intact elements containing convergently oriented and genes flanked by terminal inverted repeats (TIRs) and target site duplications with striking similarities with the corresponding elements in deuterostomes. In addition, protostome genomes contain numerous intact adjacent gene pairs that lack detectable flanking TIRs. Domains and critical active site and structural amino acids needed for endonuclease and transposase activity are present and conserved in many of the predicted RAG1L and RAG2L proteins encoded in protostome genomes.
Active transposons were present in multiple protostome lineages and many were likely transmitted vertically during protostome evolution. It appears that transposons were broadly active during bilaterian evolution, undergoing multiple duplication and loss/fossilization events, with the genes that persist in present day protostomes perhaps constituting both active transposons and domesticated genes. Our findings raise the possibility that the transposon arose earlier in evolution than previously thought, either in an early bilaterian or prior to the divergence of bilaterians and non-bilaterians, and alter our understanding of the evolutionary history of this important group of transposons.
V(D)J重组对于有颌脊椎动物的适应性免疫至关重要,且由RAG1-RAG2内切核酸酶启动。RAG1和RAG2基因被认为是从一个含有反向排列的RAG1和RAG2基因的RAG样转座子进化而来。迄今为止,仅在后口动物中令人信服地检测到了类似于这种推测的进化前体的元件,从而形成了RAG转座子最早出现在早期后口动物中的模型。
我们在原口动物的基因组中鉴定出了大量的RAG转座子,包括牡蛎和贻贝(软体动物门)以及一种纽虫(纽形动物门),以及几种刺胞动物的基因组中。系统发育分析与双侧对称动物进化枝内RAG转座子的垂直进化及其在双侧对称动物祖先中的存在一致。在原口动物中鉴定出的许多RAG转座子都是完整元件,包含反向排列的RAG1和RAG2基因,两侧是末端反向重复序列(TIR)和靶位点重复,与后口动物中的相应元件具有惊人的相似性。此外,原口动物基因组包含许多完整的RAG1-RAG2相邻基因对,这些基因对缺乏可检测到的侧翼TIR。在原口动物基因组中编码的许多预测的RAG1L和RAG2L蛋白中,存在并保守着内切核酸酶和转座酶活性所需的结构域以及关键活性位点和结构氨基酸。
活跃的RAG转座子存在于多个原口动物谱系中,并且许多可能在原口动物进化过程中垂直传播。看来RAG转座子在双侧对称动物进化过程中广泛活跃,经历了多次复制和丢失/化石形成事件,在当今原口动物中持续存在的RAG基因可能既构成活跃的RAG转座子,也构成驯化的RAG基因。我们的发现增加了RAG转座子在进化中出现的时间比以前认为的更早的可能性,要么出现在早期双侧对称动物中,要么出现在双侧对称动物和非双侧对称动物分化之前,并改变了我们对这一重要转座子群体进化历史的理解。