Unit on Genome Structure and Regulation, National Institute of Child Health and Human Development, National Institutes of Health, Bethesda, MD, USA.
Laboratory Animal Sciences Program, Frederick National Lab for Cancer Research, Frederick, MD, USA.
Nat Genet. 2023 Feb;55(2):280-290. doi: 10.1038/s41588-022-01295-6. Epub 2023 Jan 30.
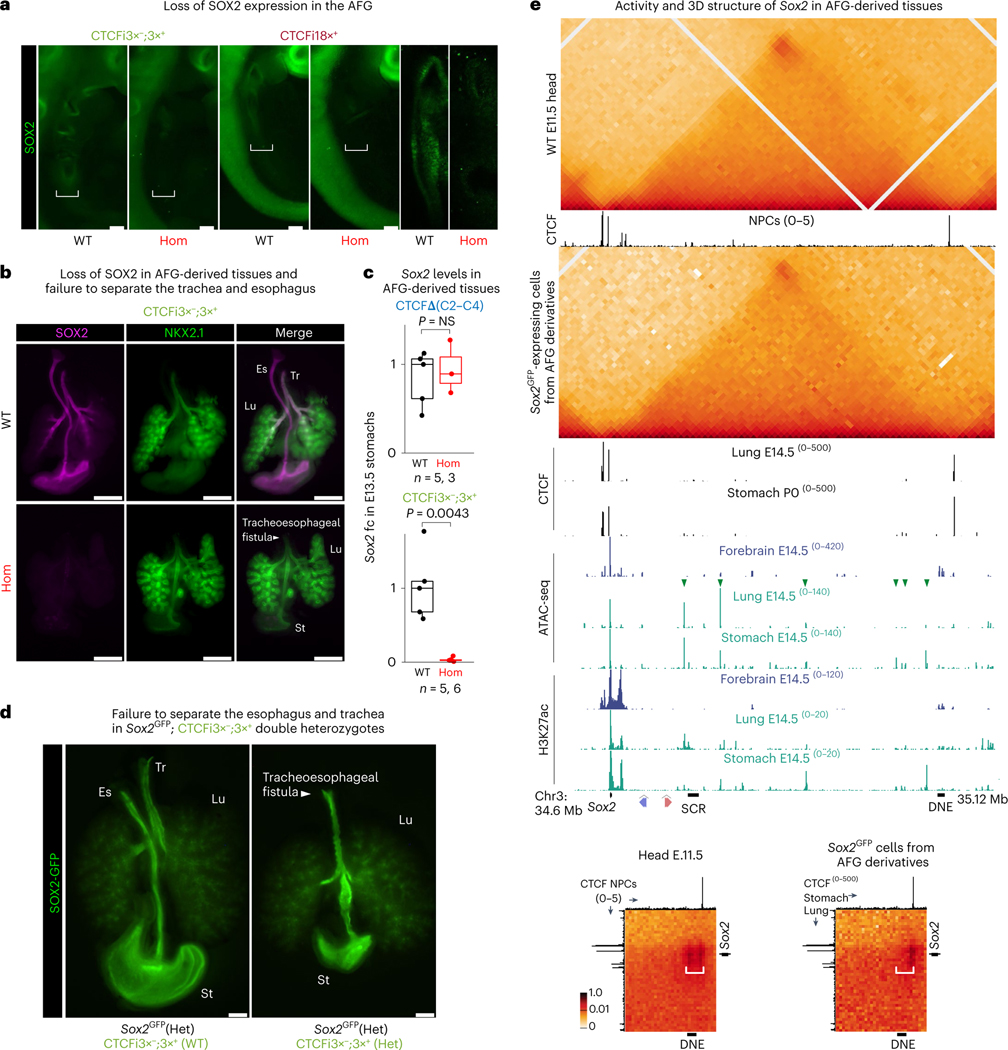
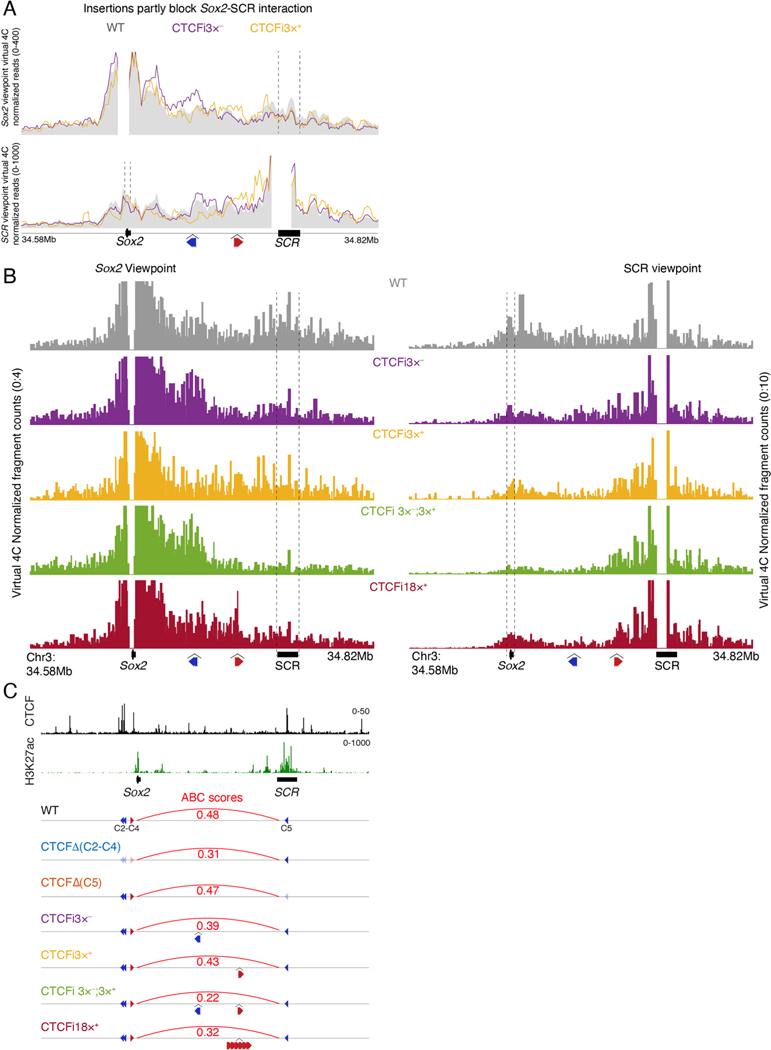
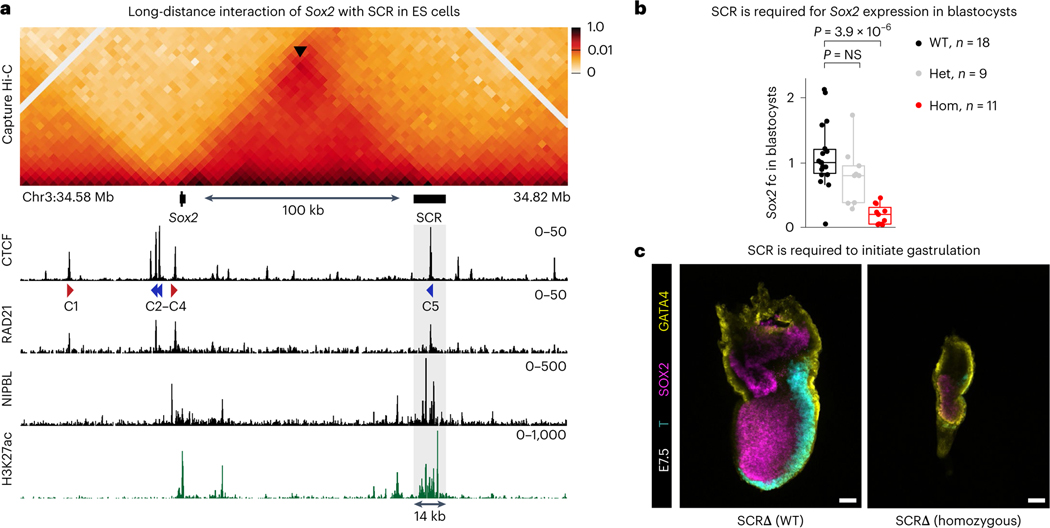
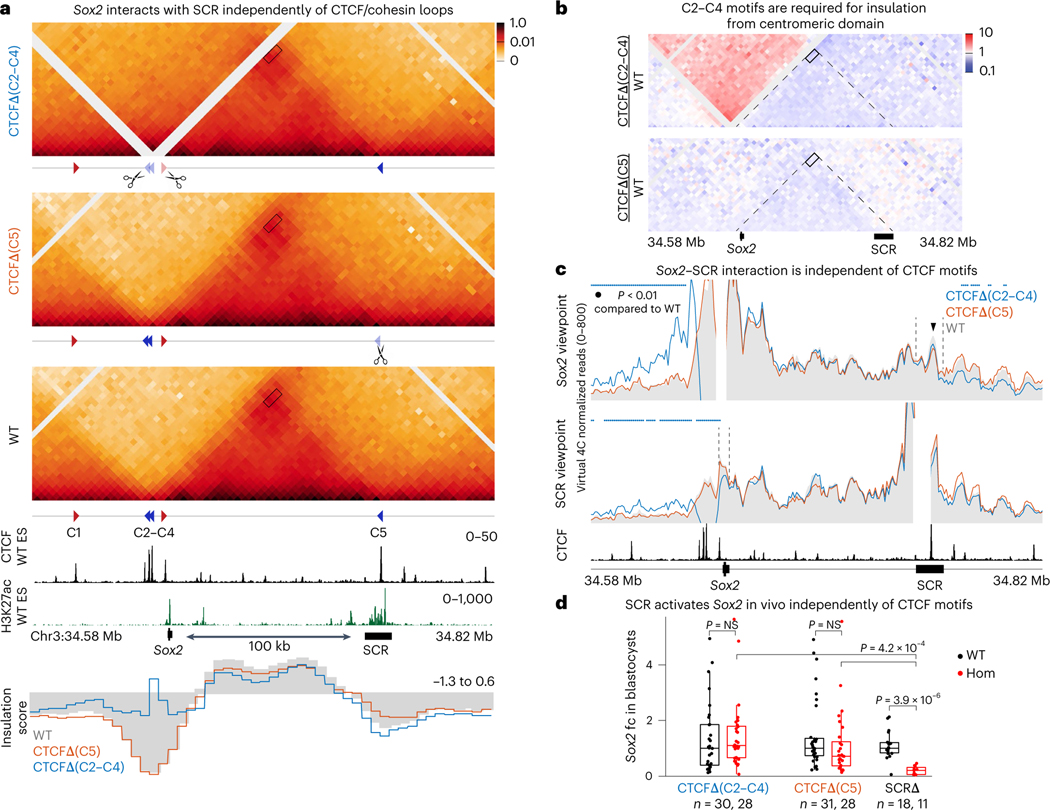
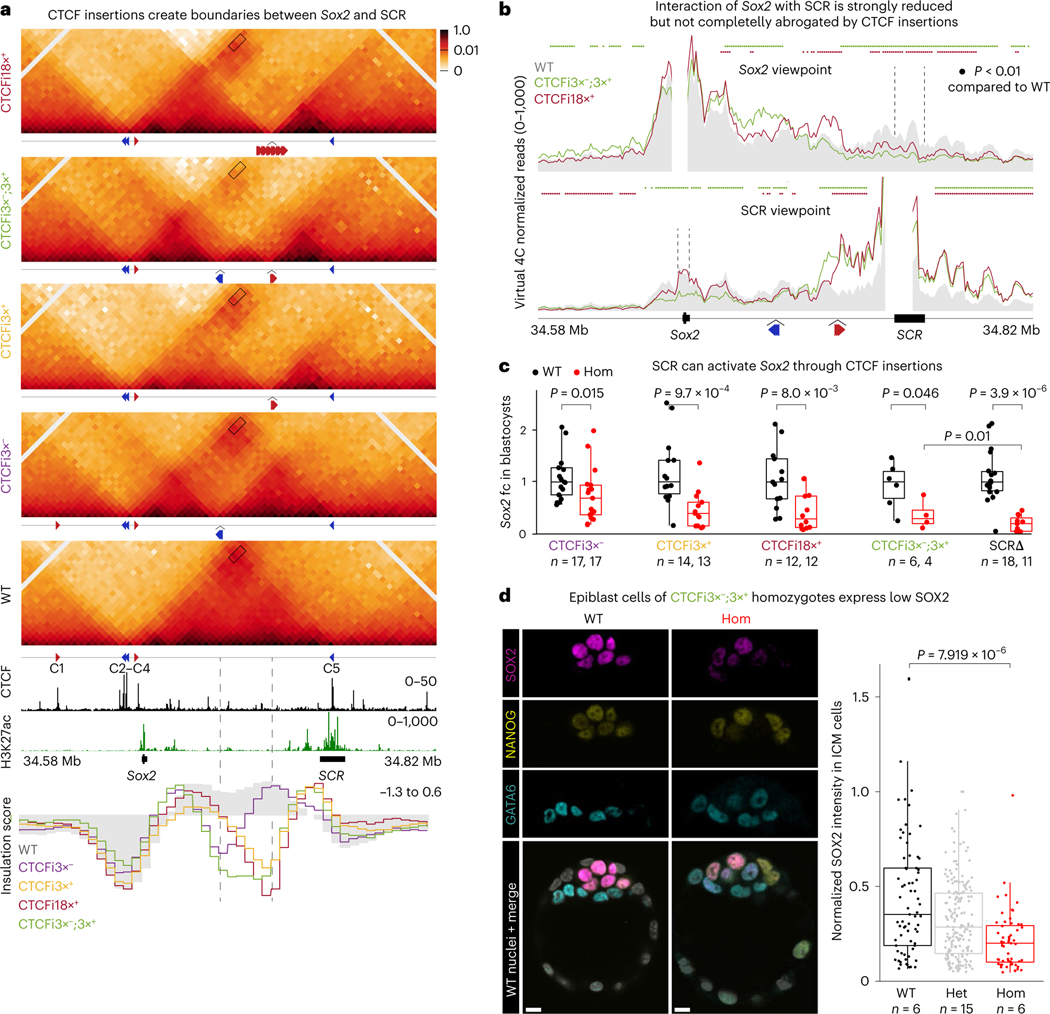
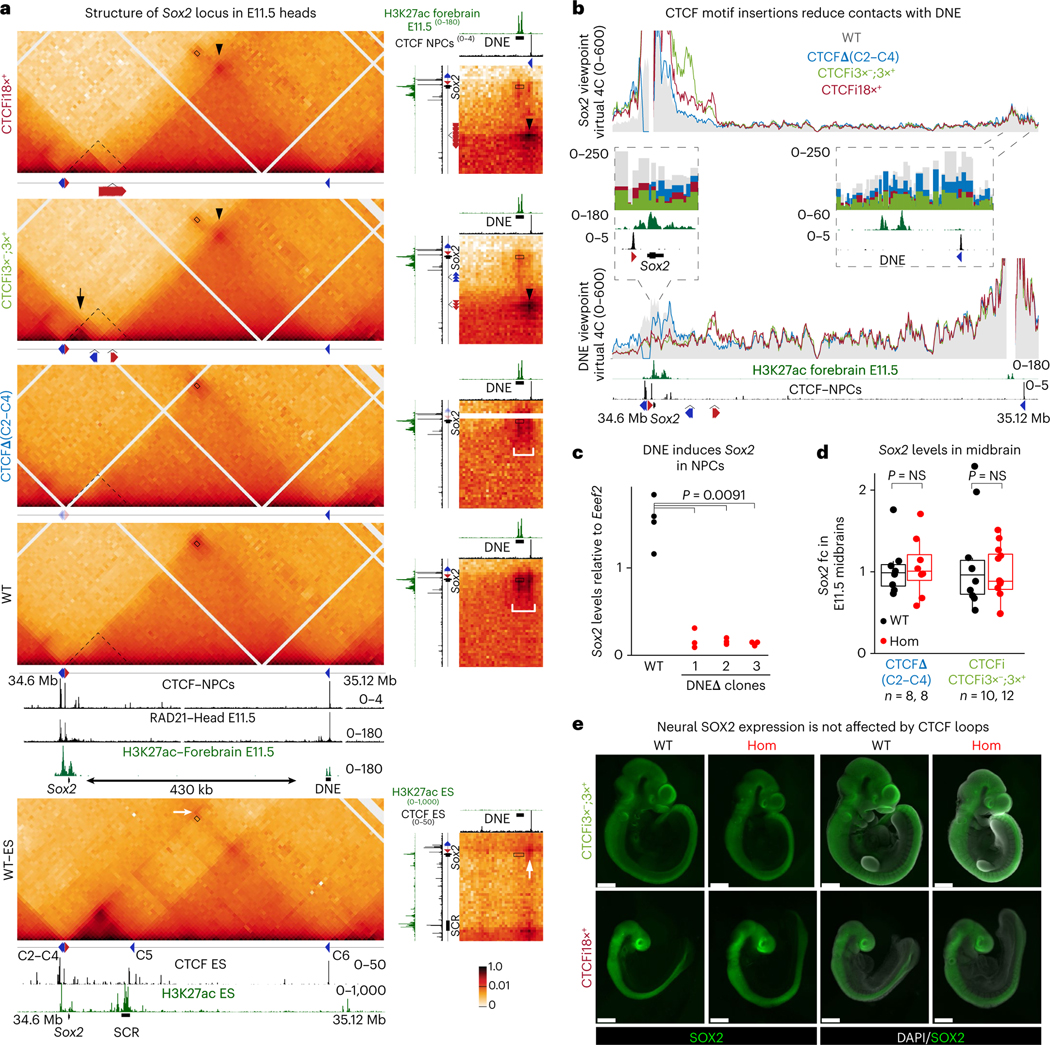
How enhancers activate their distal target promoters remains incompletely understood. Here we dissect how CTCF-mediated loops facilitate and restrict such regulatory interactions. Using an allelic series of mouse mutants, we show that CTCF is neither required for the interaction of the Sox2 gene with distal enhancers, nor for its expression. Insertion of various combinations of CTCF motifs, between Sox2 and its distal enhancers, generated boundaries with varying degrees of insulation that directly correlated with reduced transcriptional output. However, in both epiblast and neural tissues, enhancer contacts and transcriptional induction could not be fully abolished, and insertions failed to disrupt implantation and neurogenesis. In contrast, Sox2 expression was undetectable in the anterior foregut of mutants carrying the strongest boundaries, and these animals fully phenocopied loss of SOX2 in this tissue. We propose that enhancer clusters with a high density of regulatory activity can better overcome physical barriers to maintain faithful gene expression and phenotypic robustness.
增强子如何激活其远端靶启动子仍不完全清楚。在这里,我们剖析了 CTCF 介导的环如何促进和限制这种调节相互作用。使用一系列等位基因的小鼠突变体,我们表明 CTCF 既不是 Sox2 基因与远端增强子相互作用所必需的,也不是其表达所必需的。在 Sox2 和其远端增强子之间插入各种 CTCF 基序的组合,产生了具有不同程度隔离的边界,这与转录输出的减少直接相关。然而,在胚胎外胚层和神经组织中,增强子接触和转录诱导不能完全被消除,并且插入也不能破坏植入和神经发生。相比之下,携带最强边界的突变体中的前前肠中 Sox2 的表达无法检测到,并且这些动物在前肠组织中完全表现出 Sox2 的缺失。我们提出,具有高调控活性密度的增强子簇可以更好地克服物理障碍,以维持忠实的基因表达和表型稳健性。