University of California, San Francisco, San Francisco, United States.
Chan Zuckerberg Initiative, Redwood City, United States.
Elife. 2023 Feb 15;12:e81401. doi: 10.7554/eLife.81401.
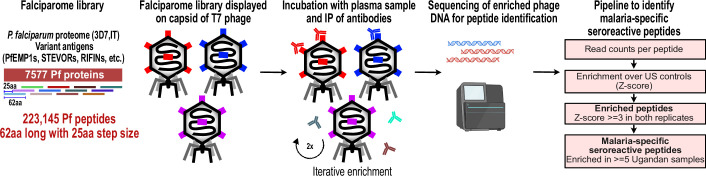
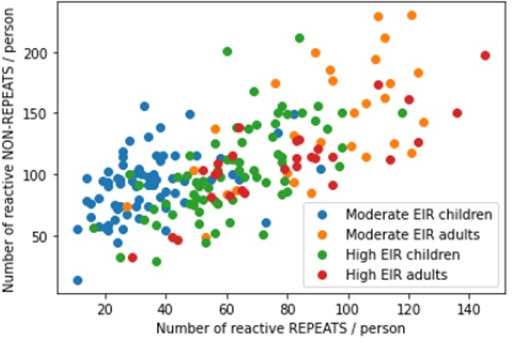
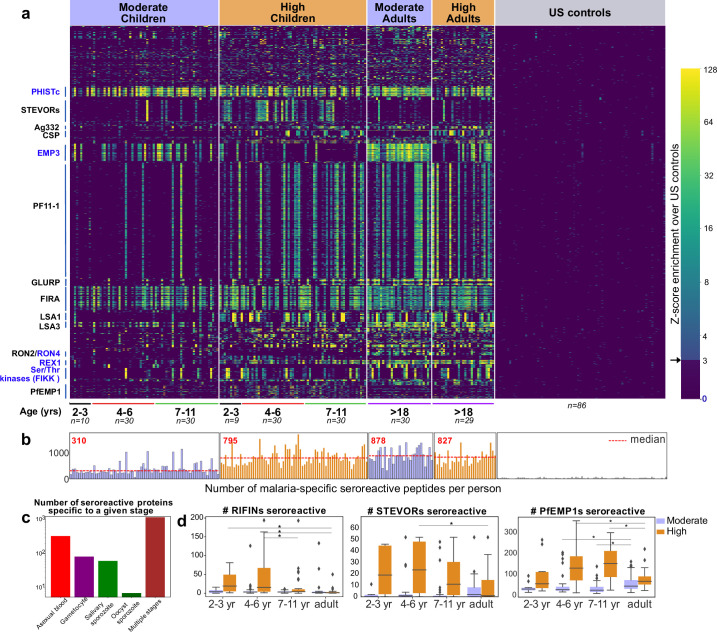
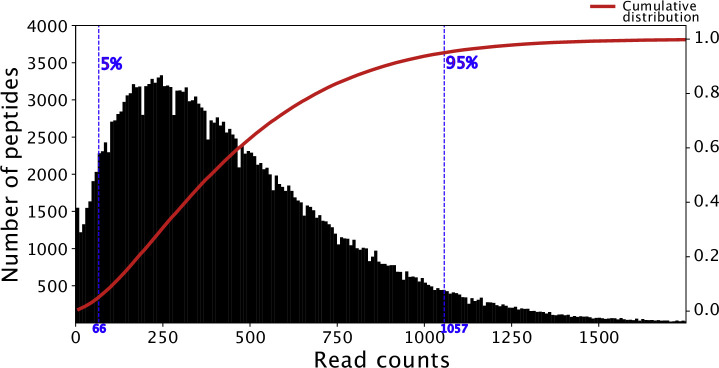
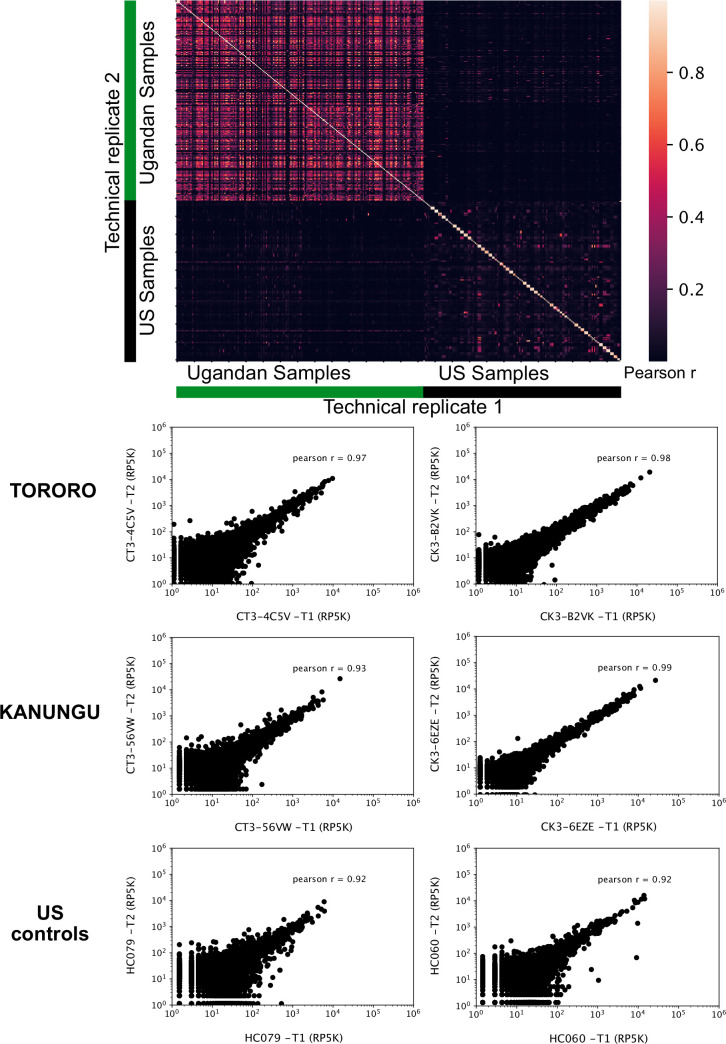
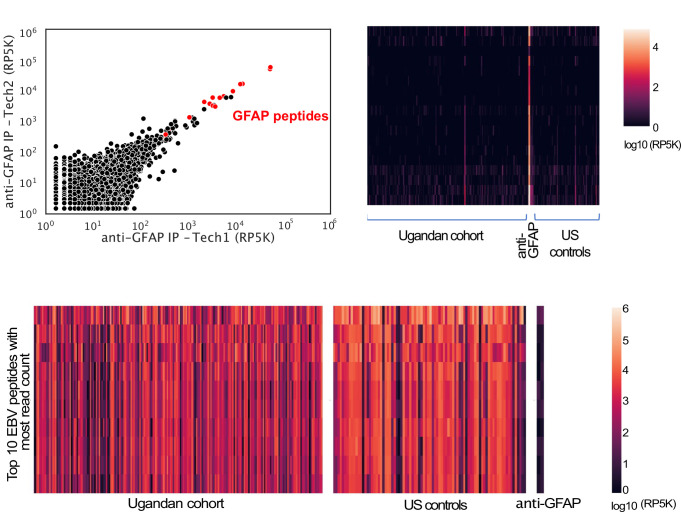
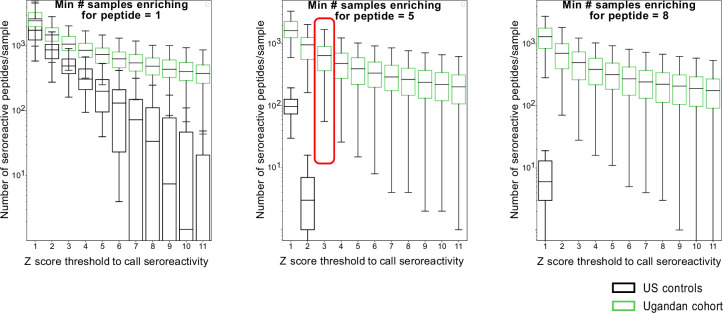
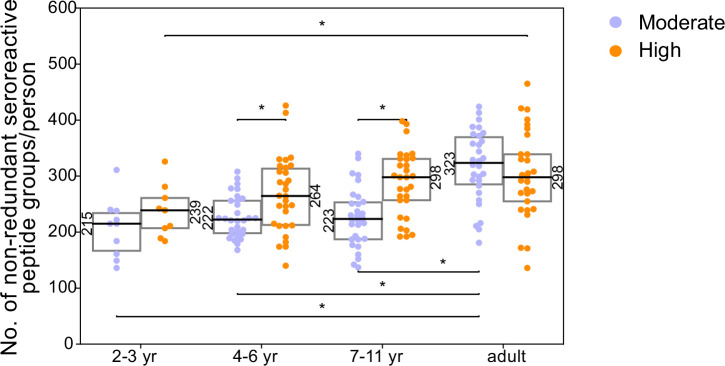
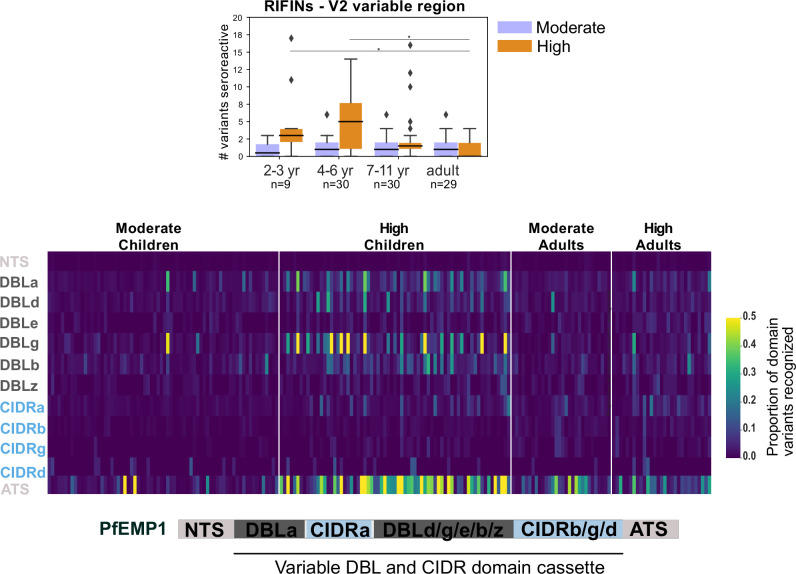
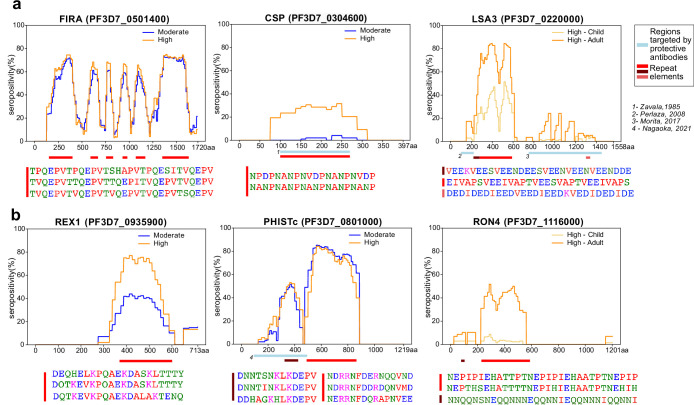
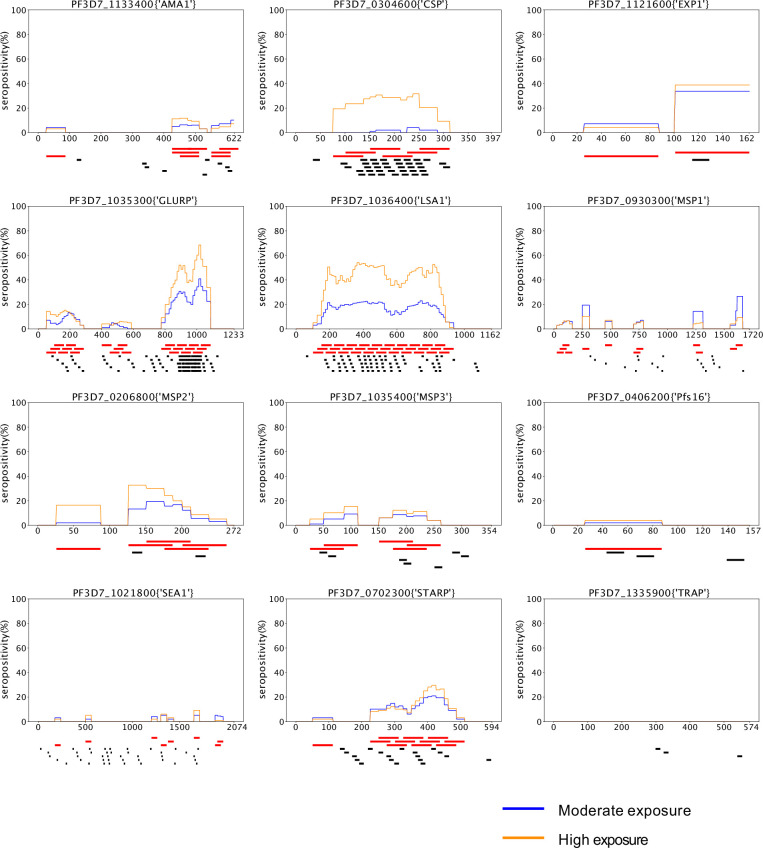
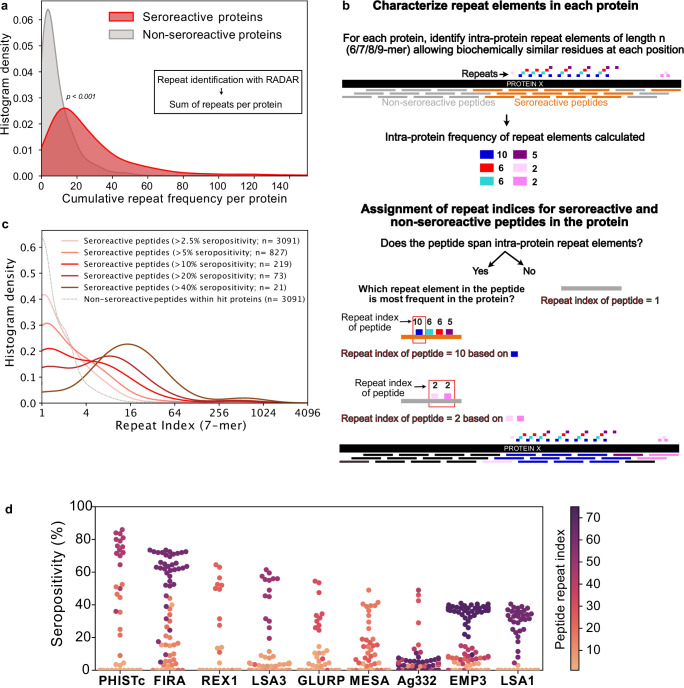
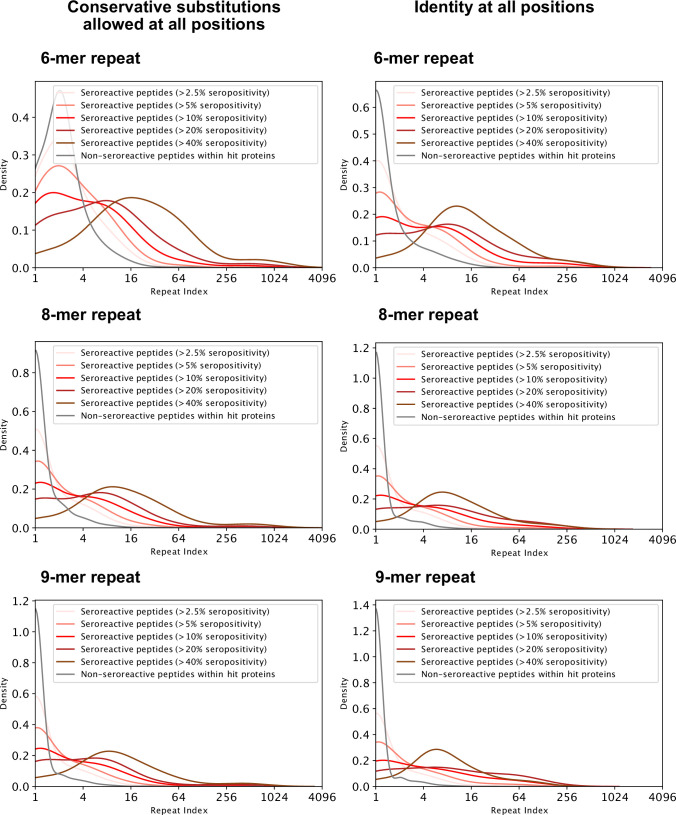
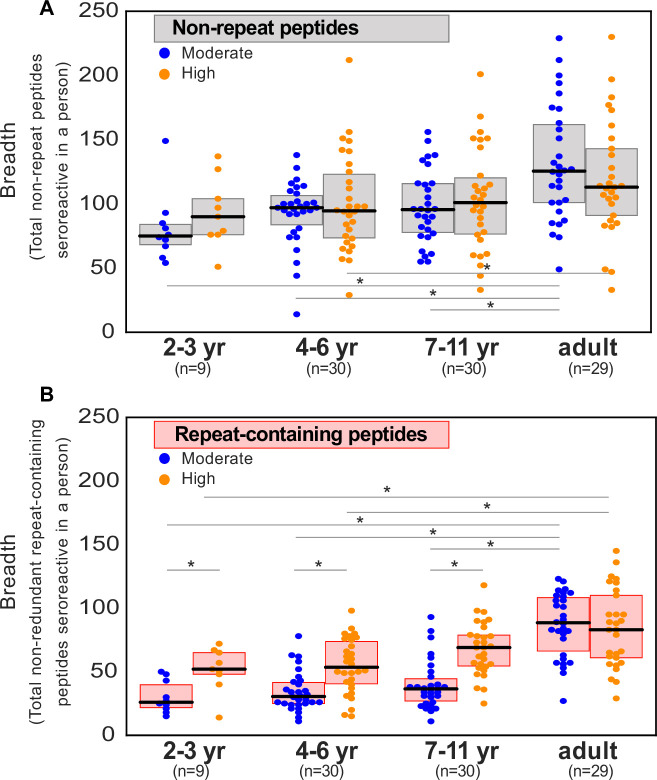
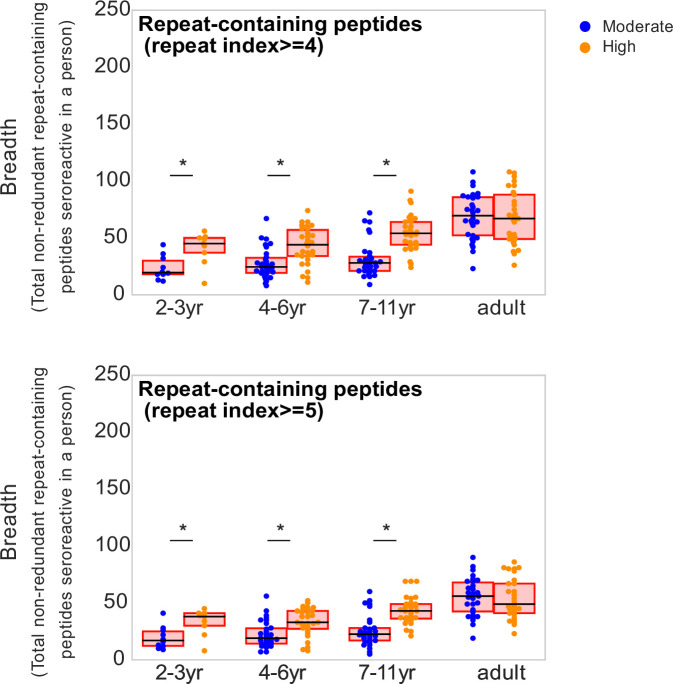
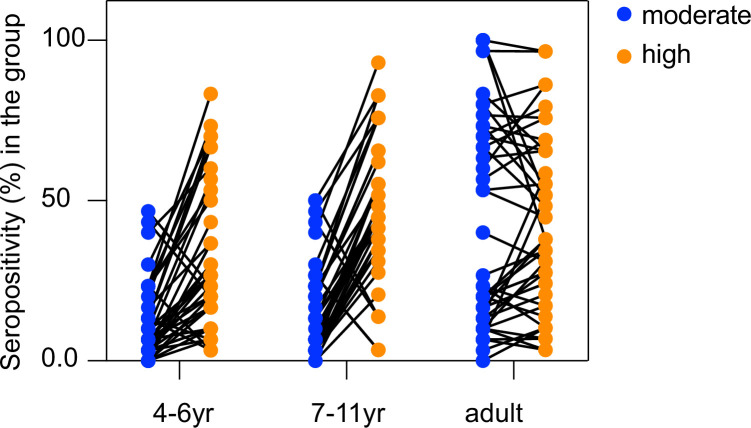
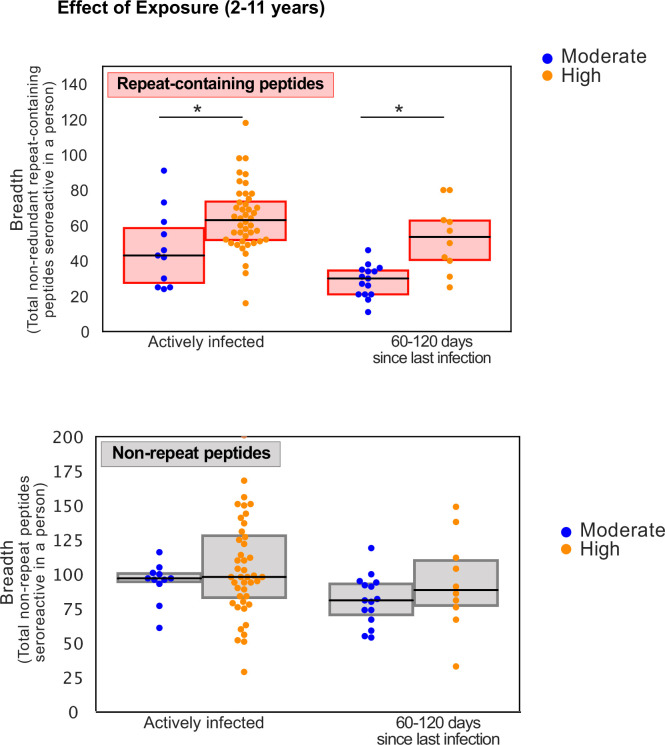
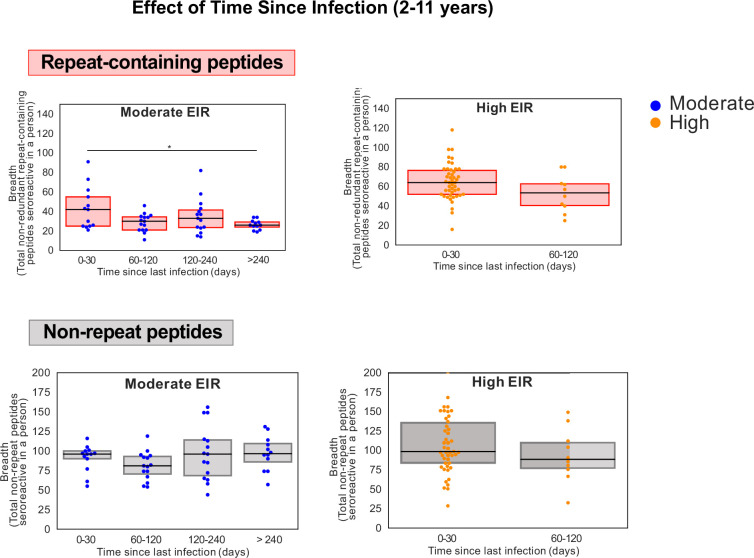
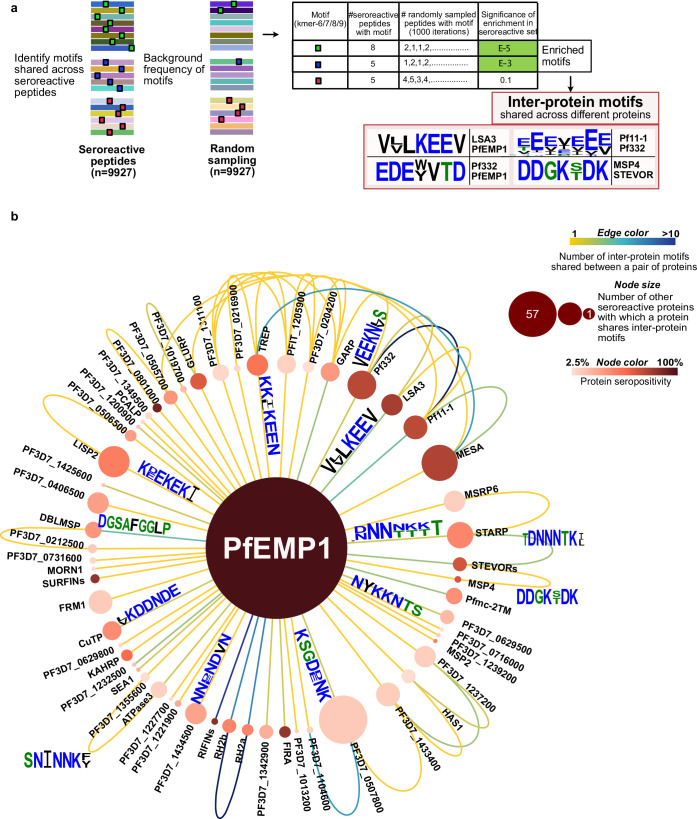
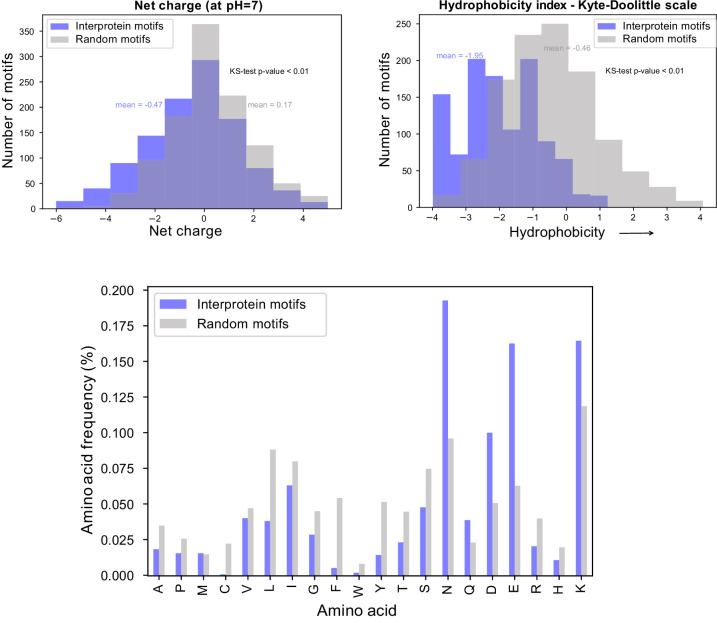
Protection against , which is primarily antibody-mediated, requires recurrent exposure to develop. The study of both naturally acquired limited immunity and vaccine induced protection against malaria remains critical for ongoing eradication efforts. Towards this goal, we deployed a customized PhIP-seq T7 phage display library containing 238,068 tiled 62-amino acid peptides, covering all known coding regions, including antigenic variants, to systematically profile antibody targets in 198 Ugandan children and adults from high and moderate transmission settings. Repeat elements - short amino acid sequences repeated within a protein - were significantly enriched in antibody targets. While breadth of responses to repeat-containing peptides was twofold higher in children living in the high versus moderate exposure setting, no such differences were observed for peptides without repeats, suggesting that antibody responses to repeat-containing regions may be more exposure dependent and/or less durable in children than responses to regions without repeats. Additionally, short motifs associated with seroreactivity were extensively shared among hundreds of antigens, potentially representing cross-reactive epitopes. PfEMP1 shared motifs with the greatest number of other antigens, partly driven by the diversity of PfEMP1 sequences. These data suggest that the large number of repeat elements and potential cross-reactive epitopes found within antigenic regions of could contribute to the inefficient nature of malaria immunity.
针对 的保护作用主要通过抗体介导,需要反复暴露才能产生。因此,研究自然获得的有限免疫力和疫苗诱导的疟疾保护作用对于正在进行的消除疟疾工作仍然至关重要。为此,我们使用定制的 PhIP-seq T7 噬菌体展示文库,该文库包含 238,068 个平铺的 62 个氨基酸肽,涵盖所有已知的编码区域,包括抗原变体,对来自高传播和中传播环境的 198 名乌干达儿童和成人的抗体靶标进行系统分析。重复元件(在蛋白质内重复的短氨基酸序列)在抗体靶标中显著富集。虽然生活在高暴露环境中的儿童对重复肽的反应广度是生活在中暴露环境中的儿童的两倍,但对于不含重复的肽则没有观察到这种差异,这表明抗体对重复区域的反应可能比没有重复的区域更依赖于暴露和/或在儿童中更持久。此外,与血清反应性相关的短基序在数百种抗原中广泛共享,这些基序可能代表交叉反应表位。PfEMP1 与其他抗原共享最多的基序,这部分是由 PfEMP1 序列的多样性驱动的。这些数据表明,抗原区域内发现的大量重复元件和潜在的交叉反应表位可能导致疟疾免疫效率低下。