Department of Genetics, University of Cambridge, Cambridge, United Kingdom.
Elife. 2023 Mar 14;12:e83364. doi: 10.7554/eLife.83364.
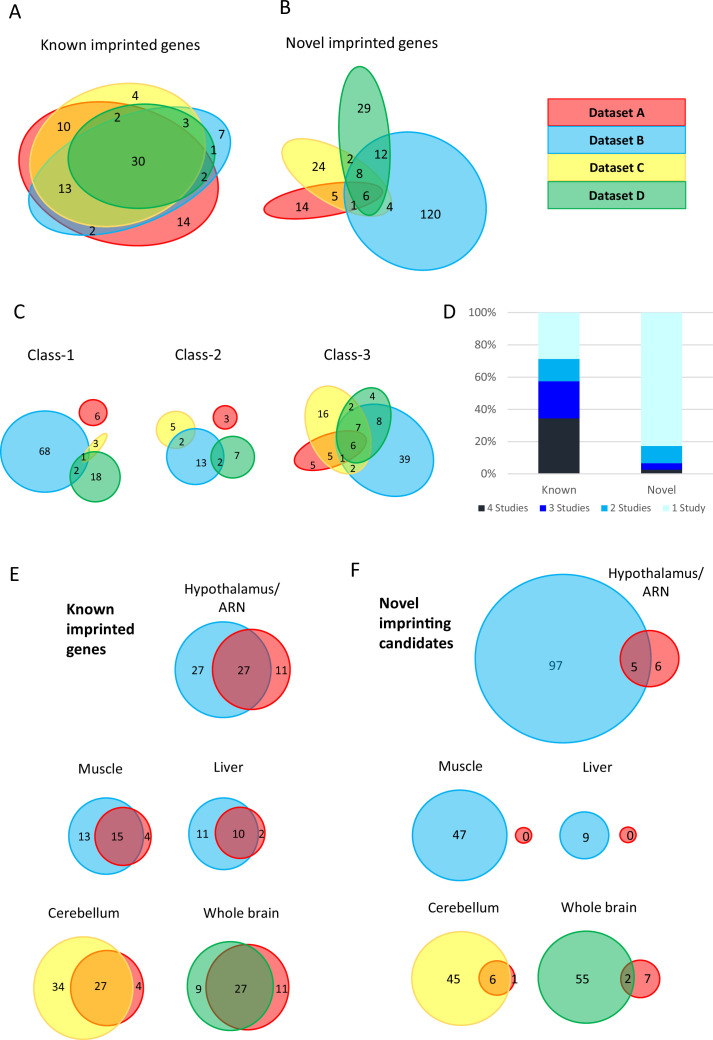
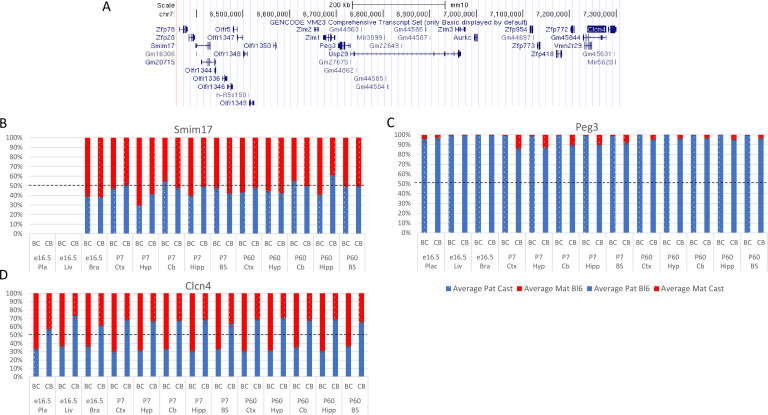
In mouse and human, genes subjected to genomic imprinting have been shown to function in development, behavior, and post-natal adaptations. Failure to correctly imprint genes in human is associated with developmental syndromes, adaptive, and metabolic disorders during life as well as numerous forms of cancer. In recent years researchers have turned to RNA-seq technologies applied to reciprocal hybrid strains of mice to identify novel imprinted genes, causing a threefold increase in genes reported as having a parental origin-specific expression bias. The functional relevance of parental origin-specific expression bias is not fully appreciated especially since many are reported with only minimal parental bias (e.g. 51:49). Here, we present an in-depth meta-analysis of previously generated RNA-seq data and show that the methods used to generate and analyze libraries greatly influence the calling of allele-specific expression. Validation experiments show that most novel genes called with parental-origin-specific allelic bias are artefactual, with the mouse strain contributing a larger effect on expression biases than parental origin. Of the weak novel genes that do validate, most are located at the periphery of known imprinted domains, suggesting they may be affected by local allele- and tissue-specific conformation. Together these findings highlight the need for robust tools, definitions, and validation of putative imprinted genes to provide meaningful information within imprinting databases and to understand the functional and mechanistic implications of the process.
在小鼠和人类中,受基因组印记调控的基因已被证明在发育、行为和出生后适应中发挥作用。人类中基因印记不正确与发育综合征、生命过程中的适应性和代谢紊乱以及许多形式的癌症有关。近年来,研究人员转向 RNA-seq 技术,应用于小鼠的正反交杂种品系,以鉴定新的印记基因,导致报告的具有亲本来源特异性表达偏倚的基因数量增加了三倍。亲本来源特异性表达偏倚的功能相关性尚未得到充分认识,特别是因为许多基因仅报告了最小的亲本偏倚(例如 51:49)。在这里,我们对之前生成的 RNA-seq 数据进行了深入的荟萃分析,结果表明,用于生成和分析文库的方法极大地影响了等位基因特异性表达的调用。验证实验表明,大多数具有亲本来源特异性等位基因偏倚的新基因都是人为的,小鼠品系对表达偏倚的影响大于亲本来源。在经过验证的少数弱效新基因中,大多数位于已知印记区域的外围,这表明它们可能受到局部等位基因和组织特异性构象的影响。这些发现共同强调了需要稳健的工具、定义和对推定的印记基因进行验证,以便在印记数据库中提供有意义的信息,并理解该过程的功能和机制影响。