The Turner Institute for Brain and Mental Health, School of Psychological Sciences and Monash Biomedical Imaging, Monash University, Clayton, Victoria, Australia.
School of Physics, University of Sydney, Camperdown, New South Wales, Australia.
Nature. 2023 Jun;618(7965):566-574. doi: 10.1038/s41586-023-06098-1. Epub 2023 May 31.
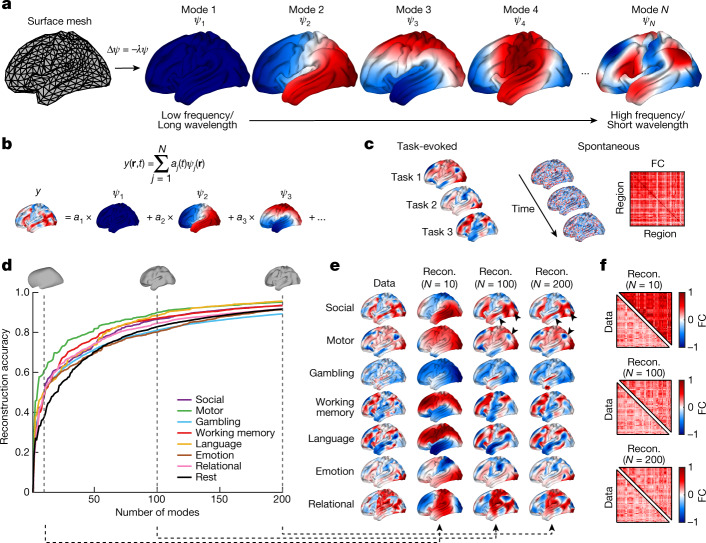
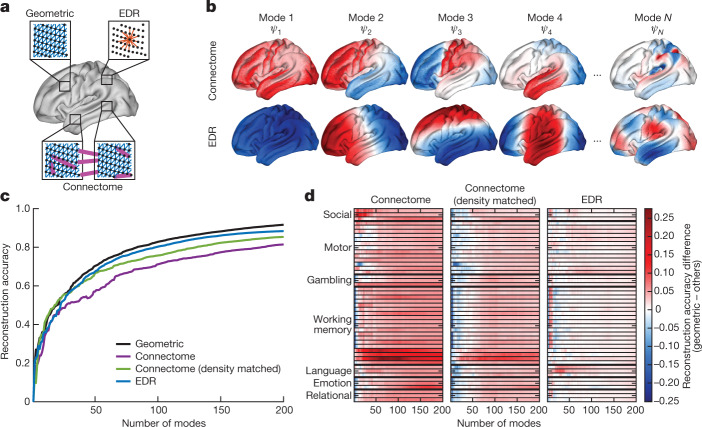
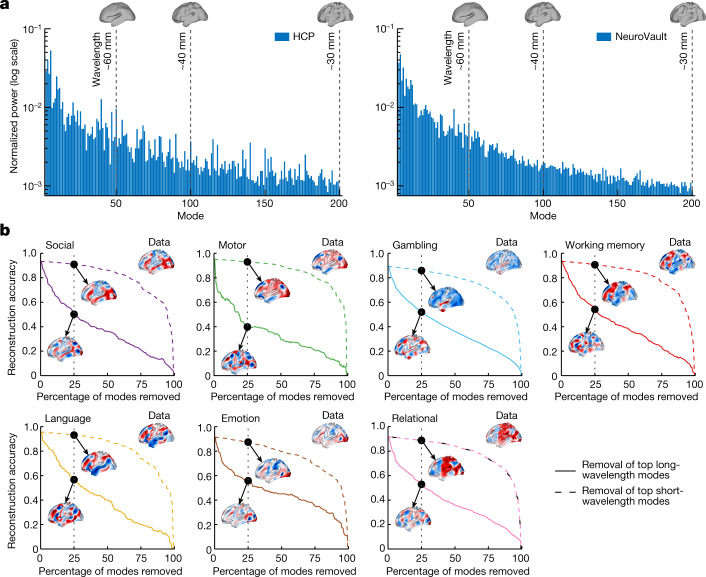
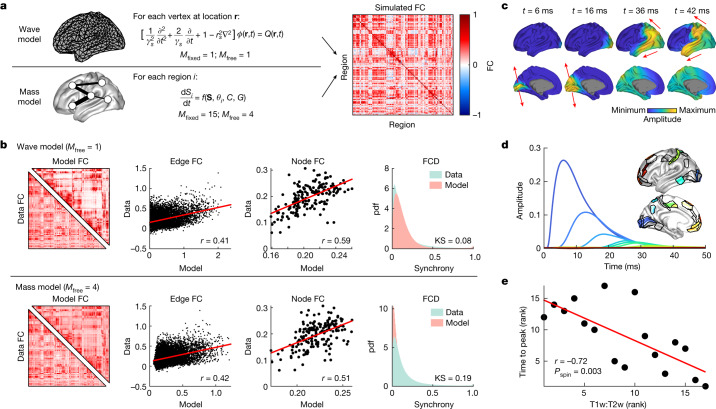
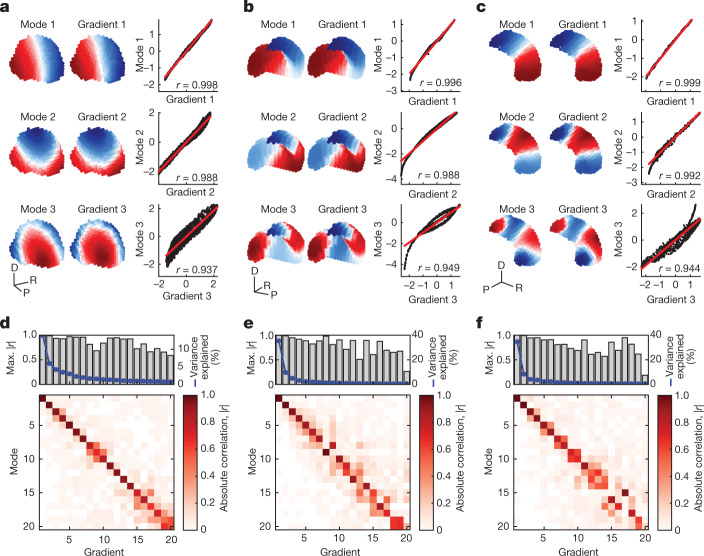
The anatomy of the brain necessarily constrains its function, but precisely how remains unclear. The classical and dominant paradigm in neuroscience is that neuronal dynamics are driven by interactions between discrete, functionally specialized cell populations connected by a complex array of axonal fibres. However, predictions from neural field theory, an established mathematical framework for modelling large-scale brain activity, suggest that the geometry of the brain may represent a more fundamental constraint on dynamics than complex interregional connectivity. Here, we confirm these theoretical predictions by analysing human magnetic resonance imaging data acquired under spontaneous and diverse task-evoked conditions. Specifically, we show that cortical and subcortical activity can be parsimoniously understood as resulting from excitations of fundamental, resonant modes of the brain's geometry (that is, its shape) rather than from modes of complex interregional connectivity, as classically assumed. We then use these geometric modes to show that task-evoked activations across over 10,000 brain maps are not confined to focal areas, as widely believed, but instead excite brain-wide modes with wavelengths spanning over 60 mm. Finally, we confirm predictions that the close link between geometry and function is explained by a dominant role for wave-like activity, showing that wave dynamics can reproduce numerous canonical spatiotemporal properties of spontaneous and evoked recordings. Our findings challenge prevailing views and identify a previously underappreciated role of geometry in shaping function, as predicted by a unifying and physically principled model of brain-wide dynamics.
大脑的解剖结构必然限制了它的功能,但具体的限制方式仍不清楚。神经科学中的经典和主导范式是,神经元的动力学是由离散的、功能专门化的细胞群体之间的相互作用驱动的,这些细胞群体通过复杂的轴突纤维网络连接。然而,神经场理论的预测,即用于模拟大规模大脑活动的成熟数学框架,表明大脑的几何形状可能是对动力学的更基本限制,而不是复杂的区域间连接。在这里,我们通过分析在自发和多种任务诱发条件下采集的人类磁共振成像数据,证实了这些理论预测。具体来说,我们表明皮质和皮质下的活动可以被简化地理解为是大脑几何形状(即形状)的基本共振模式的激发,而不是经典假设的复杂区域间连接模式的激发。然后,我们使用这些几何模式来表明,跨越超过 10000 个大脑图谱的任务诱发激活并不局限于焦点区域,这与广泛的观点相反,而是激发了具有跨越 60mm 以上波长的全脑模式。最后,我们证实了预测,即几何形状和功能之间的紧密联系是由主导的波动活动解释的,表明波动动力学可以再现自发和诱发记录的许多典型时空特性。我们的发现挑战了现有的观点,并确定了以前被低估的几何形状在塑造功能方面的作用,这与大脑整体动力学的统一和物理原则模型的预测一致。