Key Laboratory of Marine Genetics and Breeding, Ministry of Education, Ocean University of China), Qingdao, 266003, China.
BMC Genomics. 2023 Sep 18;24(1):549. doi: 10.1186/s12864-023-09639-0.
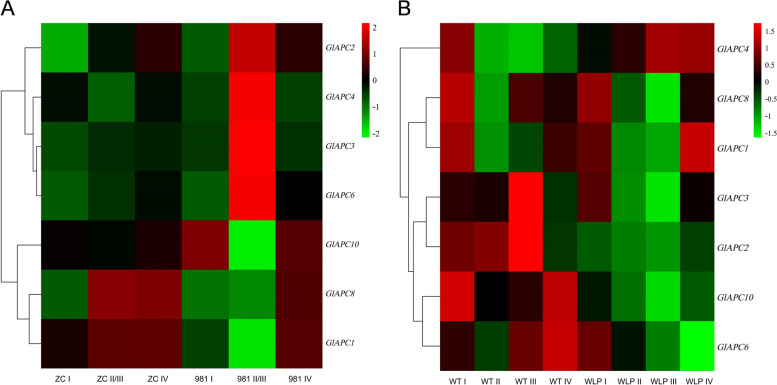
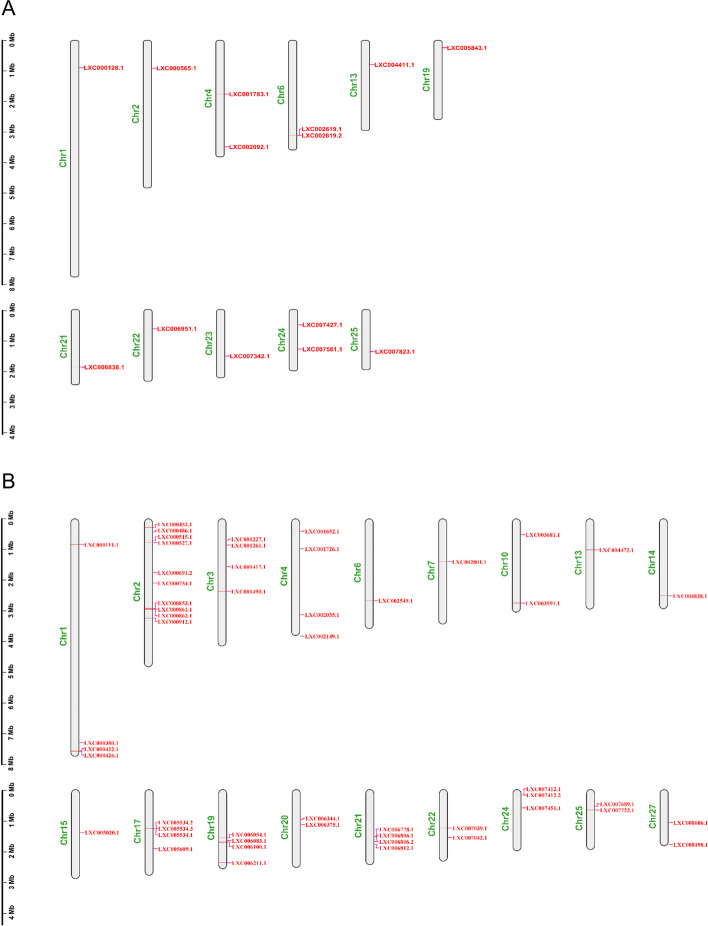
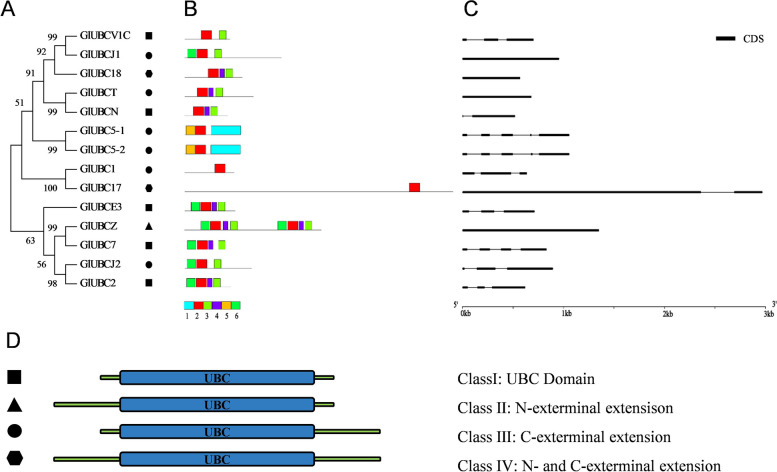
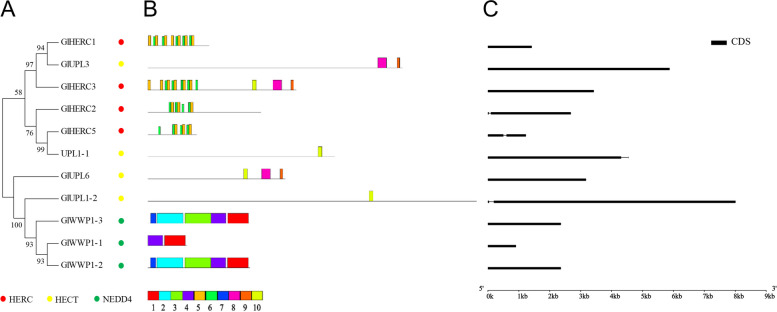
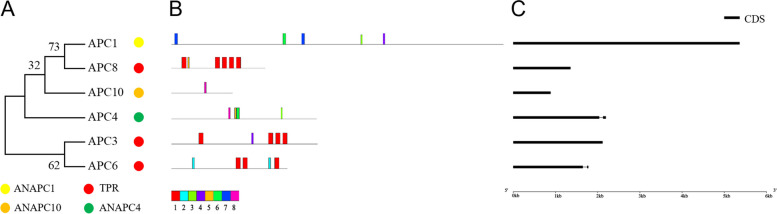
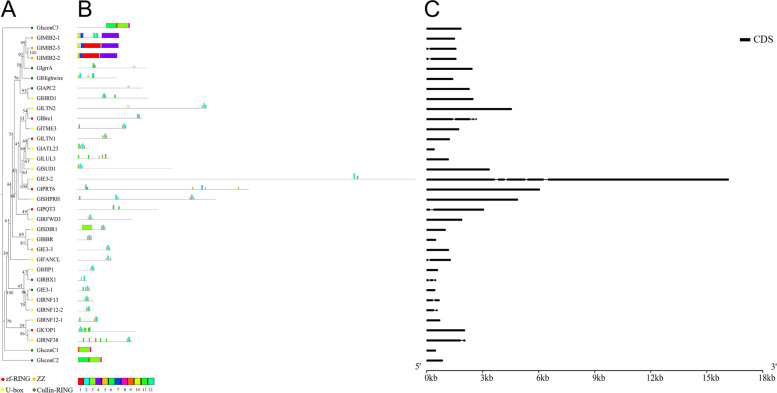
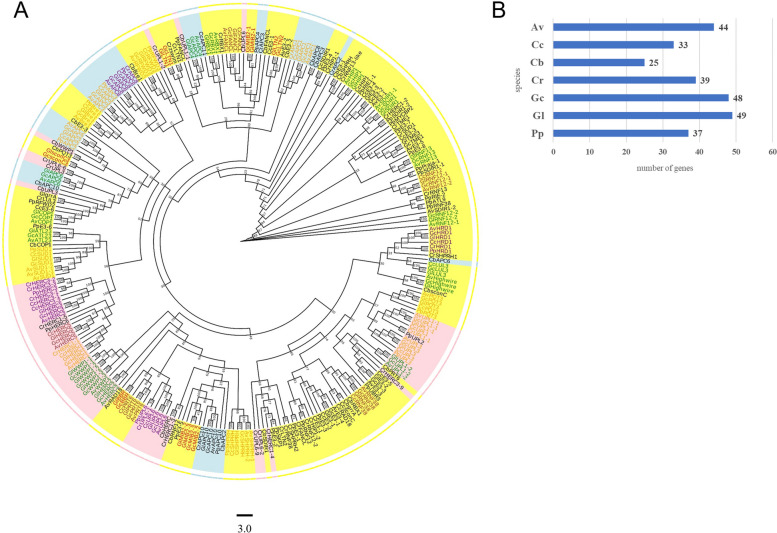
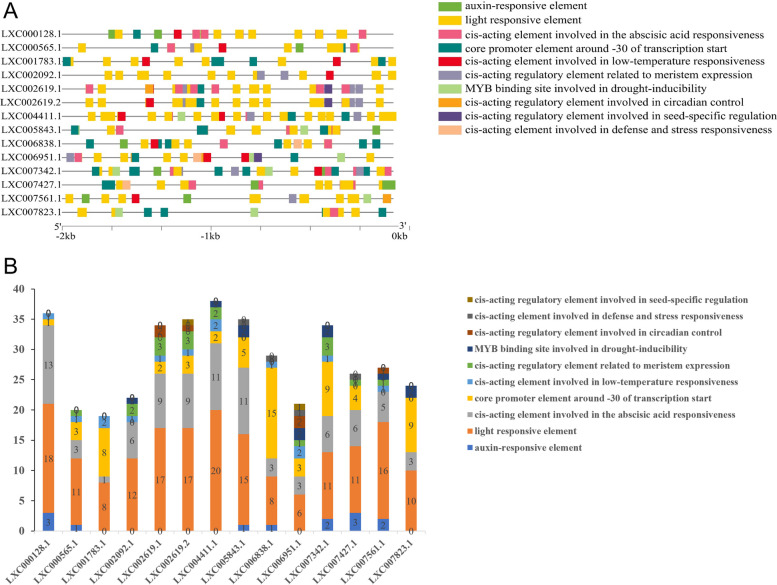
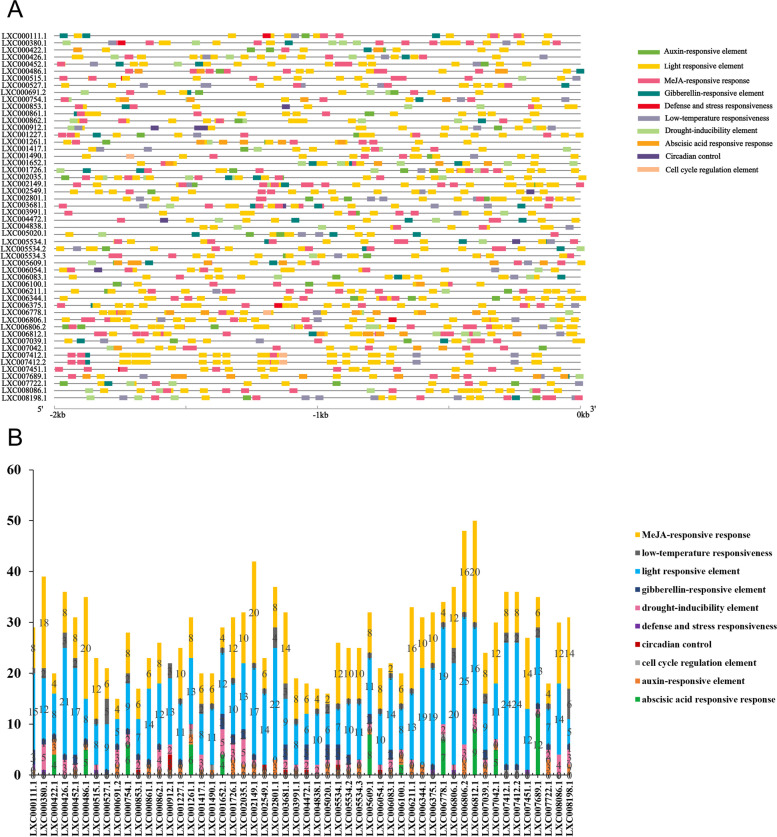
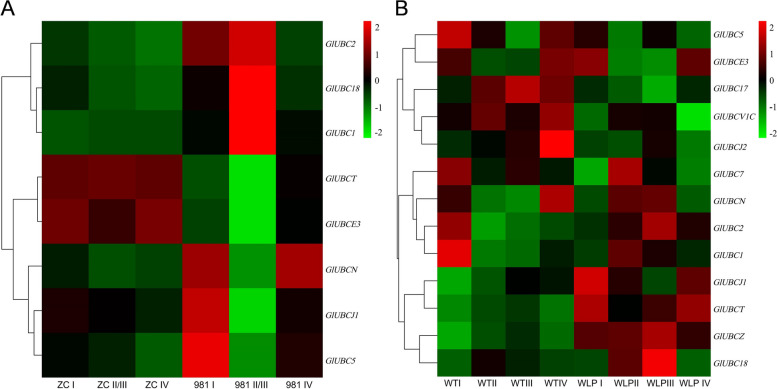
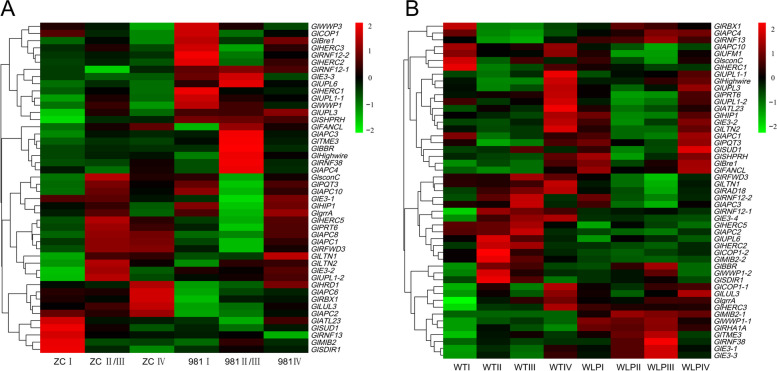
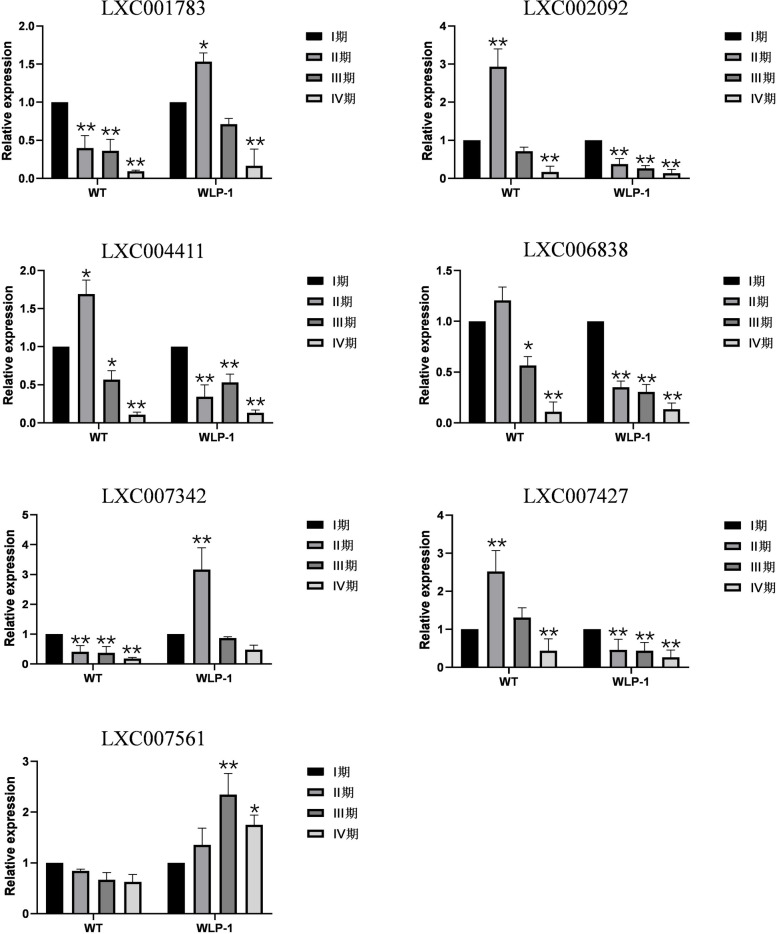
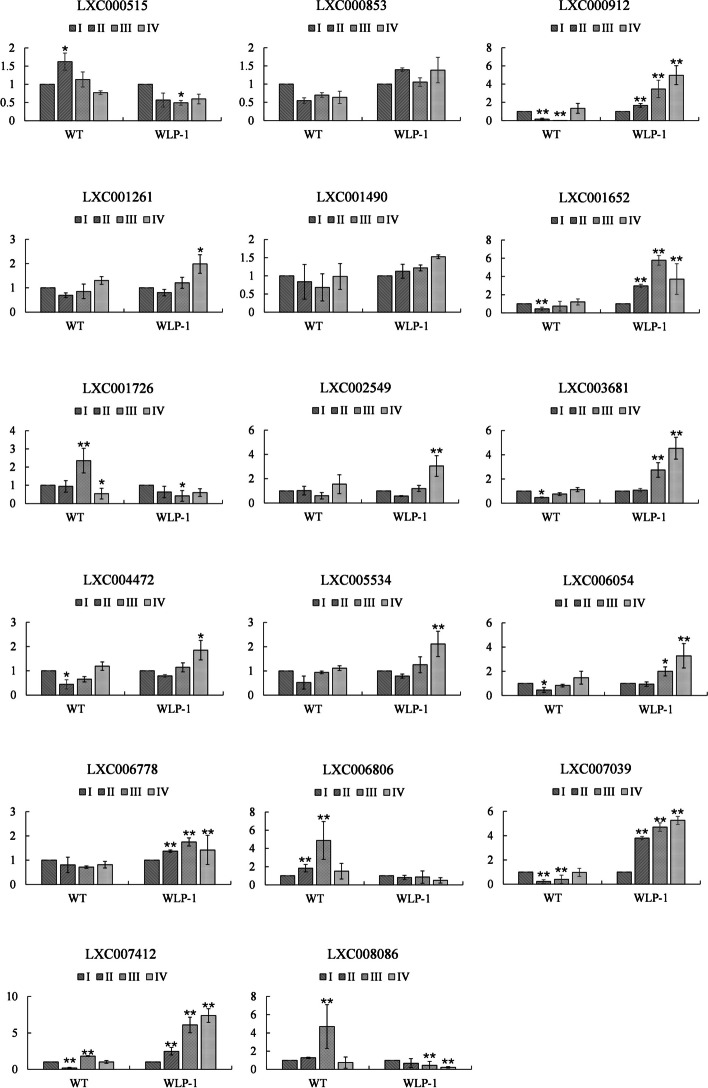
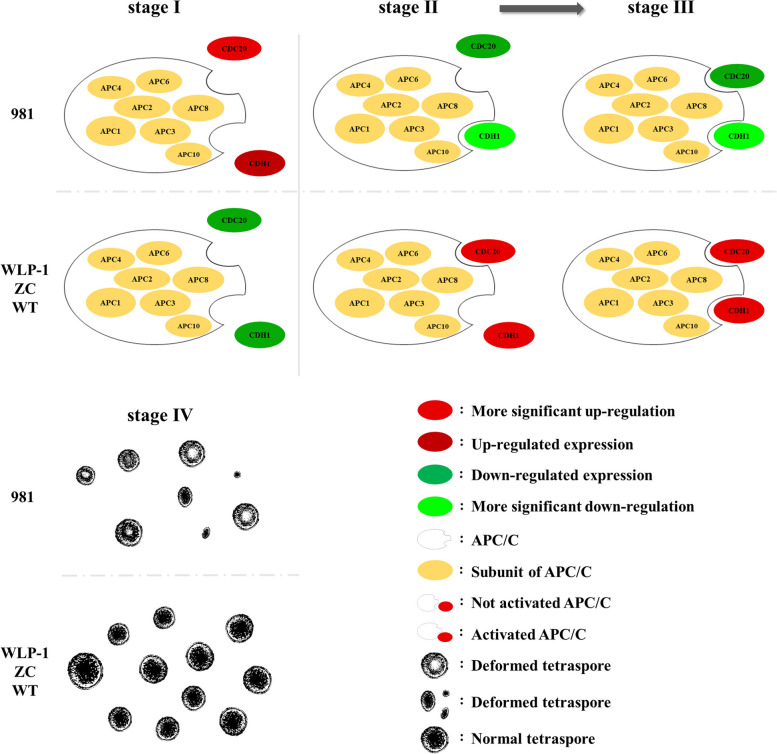
E2 ubiquitin conjugating enzymes and E3 ubiquitin ligases play important roles in the growth and development of plants and animals. To date, the systematic analysis of E2 and E3 genes in Rhodophyta is limited. In this study, 14 E2 genes and 51 E3 genes were identified in Gracilariopsis lemaneiformis, an economically important red alga. E2 genes were classified into four classes according to the structure of the conserved domain, UBC. E3 genes were classified into 12 subfamilies according to individual conserved domains. A phylogenetic tree of seven algae species showed that functional differentiation of RING-type E3s was the highest, and the similarity between orthologous genes was high except in Chlamydomonas reinhardtii and Chara braunii. RNA-seq data analysis showed significant differential expression levels of E2 and E3 genes under the life stages of tetraspore formation and release, especially GlUBCN and GlAPC3. According to GO and KEGG analysis of two transcriptomes, GlUBCN and GlAPC3 were involved in ubiquitin-mediated proteolysis, and other subunits of the anaphase promoting complex or cyclosome (APC/C) and its activators GlCDC20 and GlCDH1 were also enriched into this process. The CDH1 and CDC20 in 981 were down-regulated during tetraspores formation and release, with the down-regulation of CDH1 being particularly significant; CDH1 and CDC20 in WLP-1, ZC, and WT were up-regulated during tetraspores formation and release, with CDC20 being more significantly up-regulated. Therefore, GlCDH1, rather than GlCDC20, in '981' might play the leading role in the activation of the APC/C, and GlCDC20 might play the leading role rather than GlCDH1 in strains WLP-1, ZC and wild type. The low fertility of cultivar 981 might be highly correlated with the inactivity of activators CDH1 and CDC20. This study provided a basic and comprehensive understanding of characteristic of E2 and E3 genes in Gp. lemaneiformis and set a foundation for further understanding of E2 ubiquitin conjugating enzymes and E3 ubiquitin ligase in regulating tetrasporophytes development of Gp. lemaneiformis.
E2 泛素连接酶和 E3 泛素连接酶在动植物的生长和发育中发挥着重要作用。迄今为止,对红藻门中 E2 和 E3 基因的系统分析是有限的。在这项研究中,从经济上重要的红藻龙须菜中鉴定出了 14 个 E2 基因和 51 个 E3 基因。根据保守结构域 UBC,E2 基因被分为四个类。根据单个保守结构域,E3 基因被分为 12 个亚家族。七种藻类的系统发育树表明,RING 型 E3s 的功能分化最高,除了莱茵衣藻和轮藻外,同源基因之间的相似性很高。RNA-seq 数据分析表明,在四分孢子形成和释放的生命阶段,E2 和 E3 基因的表达水平存在显著差异,尤其是 GlUBCN 和 GlAPC3。根据两个转录组的 GO 和 KEGG 分析,GlUBCN 和 GlAPC3 参与了泛素介导的蛋白质水解,其他有丝分裂促进复合物或细胞周期蛋白(APC/C)及其激活剂 GlCDC20 和 GlCDH1 的亚基也被富集到这个过程中。在四分孢子形成和释放过程中,981 中的 CDH1 和 CDC20 下调,其中 CDH1 的下调尤为显著;在 WLP-1、ZC 和 WT 中,CDH1 和 CDC20 在四分孢子形成和释放过程中上调,其中 CDC20 上调更为显著。因此,在 '981' 中,GlCDH1 而不是 GlCDC20 可能在 APC/C 的激活中起主导作用,而 GlCDC20 可能在 WLP-1、ZC 和野生型中起主导作用,而不是 GlCDH1。栽培品种 981 低育性可能与激活剂 CDH1 和 CDC20 的不活跃高度相关。本研究为龙须菜 E2 和 E3 基因的特征提供了一个基本而全面的了解,并为进一步了解 E2 泛素连接酶和 E3 泛素连接酶在调节龙须菜四分孢子体发育中的作用奠定了基础。