Venkatachalam Thejasvi, Mannimala Sushma, Soto Martha C
Department of Pathology and Laboratory Medicine, Rutgers - Robert Wood Johnson Medical School, Piscataway, NJ, USA.
bioRxiv. 2023 Oct 5:2023.10.04.560868. doi: 10.1101/2023.10.04.560868.
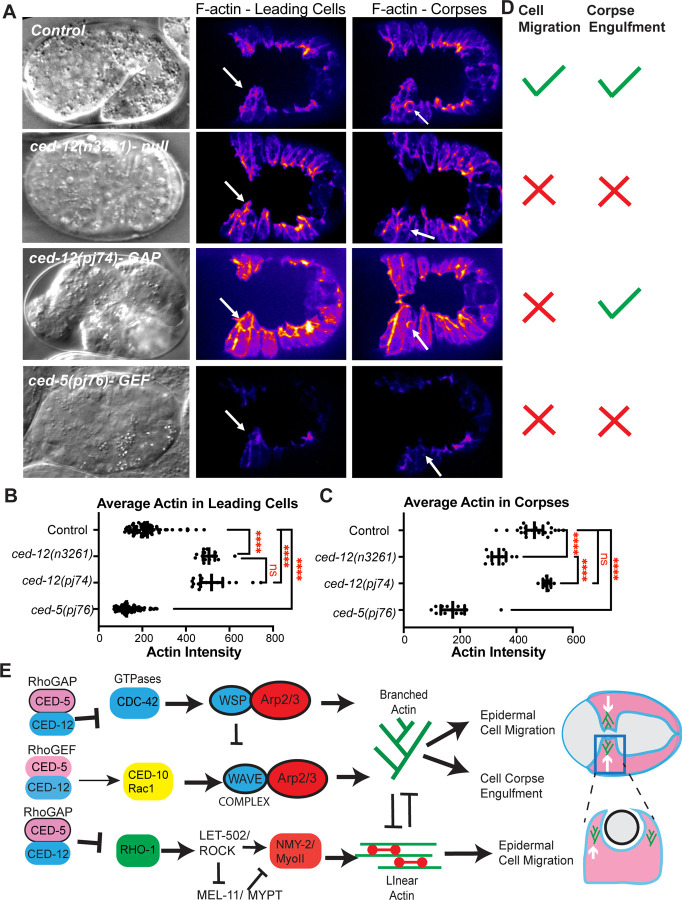
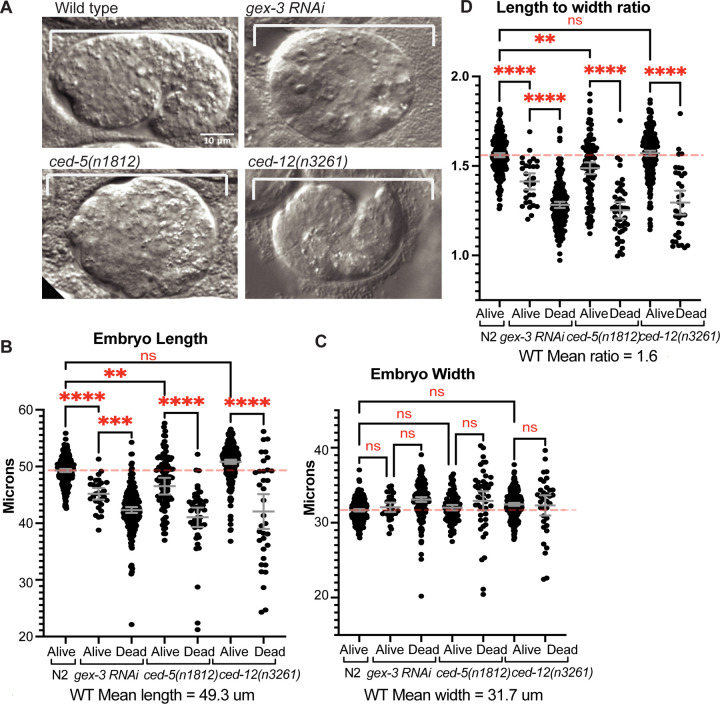
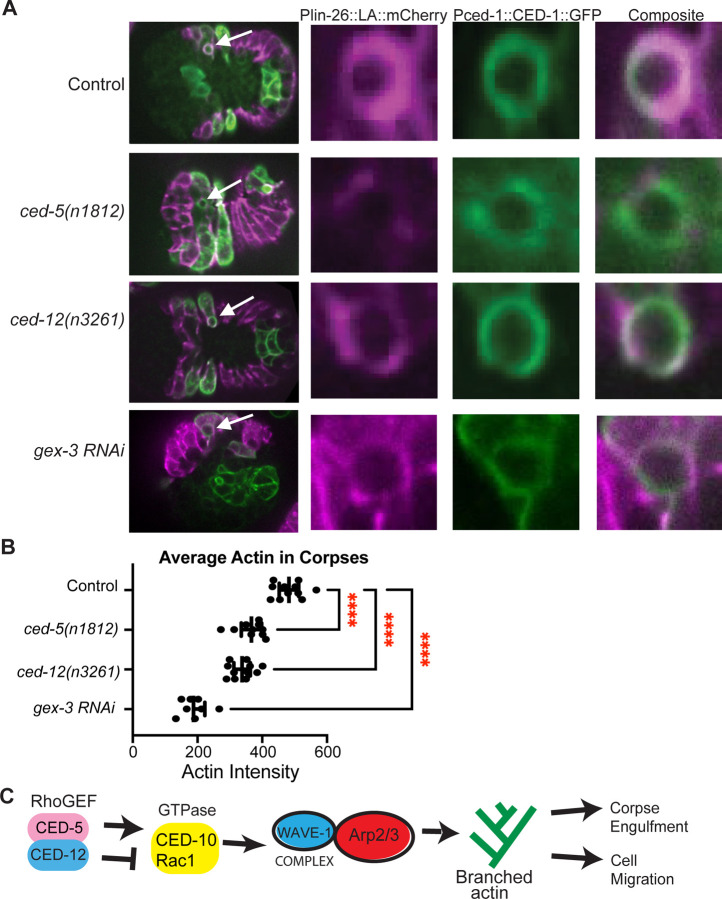
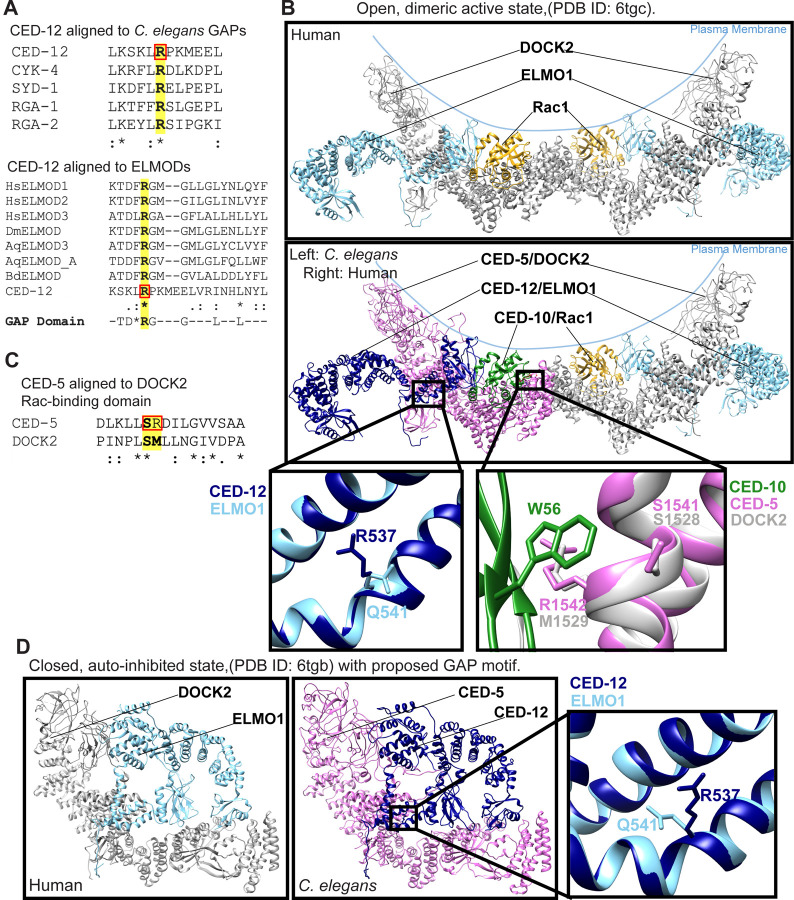
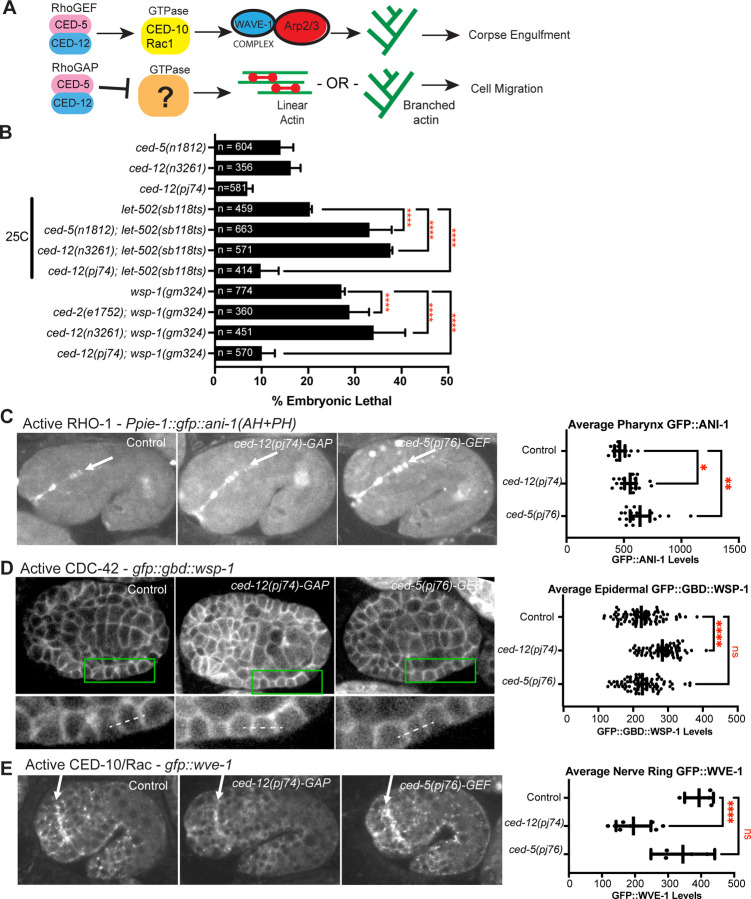
Coordinated activation and inhibition of F-actin supports the movements of morphogenesis. Understanding the proteins that regulate F-actin is important, since these proteins are mis-regulated in diseases like cancer. Our studies of embryonic epidermal morphogenesis identified the GTPase CED-10/Rac1 as an essential activator of F-actin. However, we need to identify the GEF, or Guanine-nucleotide Exchange Factor, that activates CED-10/Rac1 during embryonic cell migrations. The two-component GEF, CED-5/CED-12, is known to activate CED-10/Rac1 to promote cell movements that result in the engulfment of dying cells during embryogenesis, and a later cell migration of the larval Distal Tip Cell. It is believed that CED-5/CED-12 powers cellular movements of corpse engulfment and DTC migration by promoting F-actin formation. Therefore, we tested if CED-5/CED-12 was involved in embryonic migrations, and got a contradictory result. CED-5/CED-12 definitely support embryonic migrations, since their loss led to embryos that died due to failed epidermal cell migrations. However, CED-5/CED-12 inhibited F-actin in the migrating epidermis, the opposite of what was expected for a CED-10 GEF. To address how CED-12/CED-5 could have two opposing effects on F-actin, during corpse engulfment and cell migration, we investigated if CED-12 harbors GAP (GTPase Activating Protein) functions. A candidate GAP region in CED-12 faces away from the CED-5 GEF catalytic region. Mutating a candidate catalytic Arginine in the CED-12 GAP region (R537A) altered the epidermal cell migration function, and not the corpse engulfment function. A candidate GEF region on CED-5 faces towards Rac1/CED-10. Mutating Serine-Arginine in CED-5/DOCK predicted to bind and stabilize Rac1 for catalysis, resulted in loss of both ventral enclosure and corpse engulfment. Genetic and expression studies showed the GEF and GAP functions act on different GTPases. Thus, we propose CED-5/CED-12 support the cycling of multiple GTPases, by using distinct domains, to both promote and inhibit F-actin nucleation.
F-肌动蛋白的协同激活与抑制支持形态发生运动。了解调节F-肌动蛋白的蛋白质很重要,因为这些蛋白质在癌症等疾病中会出现调节异常。我们对胚胎表皮形态发生的研究确定GTP酶CED-10/Rac1是F-肌动蛋白的一种重要激活剂。然而,我们需要确定在胚胎细胞迁移过程中激活CED-10/Rac1的鸟嘌呤核苷酸交换因子(GEF)。已知双组分GEF,即CED-5/CED-12,可激活CED-10/Rac1,以促进细胞运动,这些运动导致胚胎发育过程中死亡细胞的吞噬,以及幼虫远端顶端细胞随后的细胞迁移。据信,CED-5/CED-12通过促进F-肌动蛋白的形成来推动尸体吞噬和远端顶端细胞迁移的细胞运动。因此,我们测试了CED-5/CED-12是否参与胚胎迁移,结果却相互矛盾。CED-5/CED-12肯定支持胚胎迁移,因为它们的缺失会导致胚胎因表皮细胞迁移失败而死亡。然而,CED-5/CED-12在迁移的表皮中抑制F-肌动蛋白,这与对CED-10 GEF的预期相反。为了解决CED-12/CED-5如何在尸体吞噬和细胞迁移过程中对F-肌动蛋白产生两种相反作用的问题,我们研究了CED-12是否具有GTP酶激活蛋白(GAP)功能。CED-12中的一个候选GAP区域背对CED-5 GEF催化区域。在CED-12 GAP区域(R537A)中突变一个候选催化精氨酸会改变表皮细胞迁移功能,但不会改变尸体吞噬功能。CED-5上的一个候选GEF区域朝向Rac1/CED-10。在CED-5/DOCK中突变预测会结合并稳定Rac1以进行催化的丝氨酸-精氨酸,会导致腹侧包被和尸体吞噬功能丧失。遗传和表达研究表明,GEF和GAP功能作用于不同的GTP酶。因此,我们提出CED-5/CED-12通过使用不同的结构域来支持多种GTP酶的循环,以促进和抑制F-肌动蛋白成核。