Department of Molecular and Cellular Physiology, Stanford University, Stanford, United States.
Department of Chemistry, Stanford University, Stanford, United States.
Elife. 2024 Feb 12;12:RP90648. doi: 10.7554/eLife.90648.
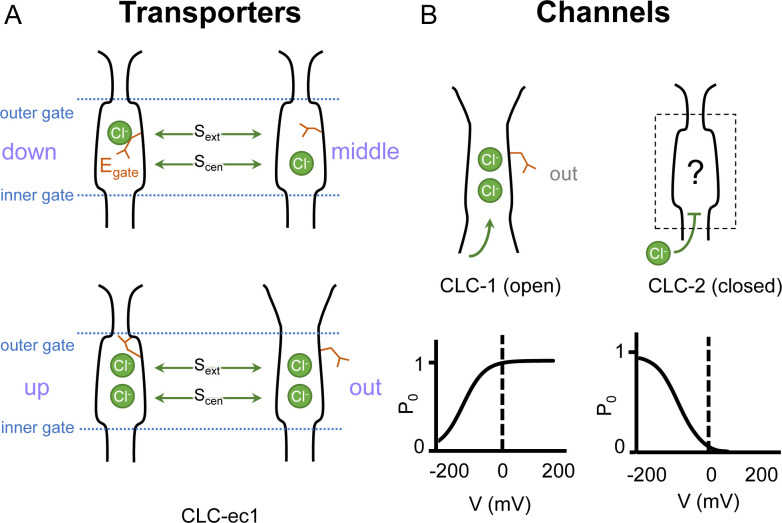
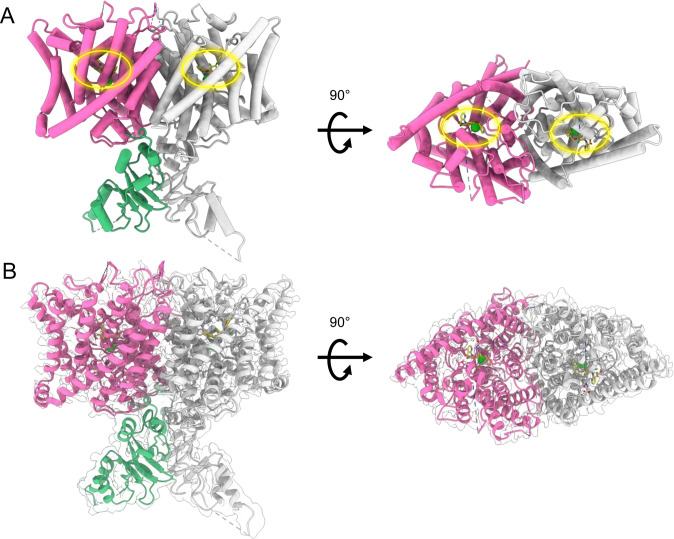
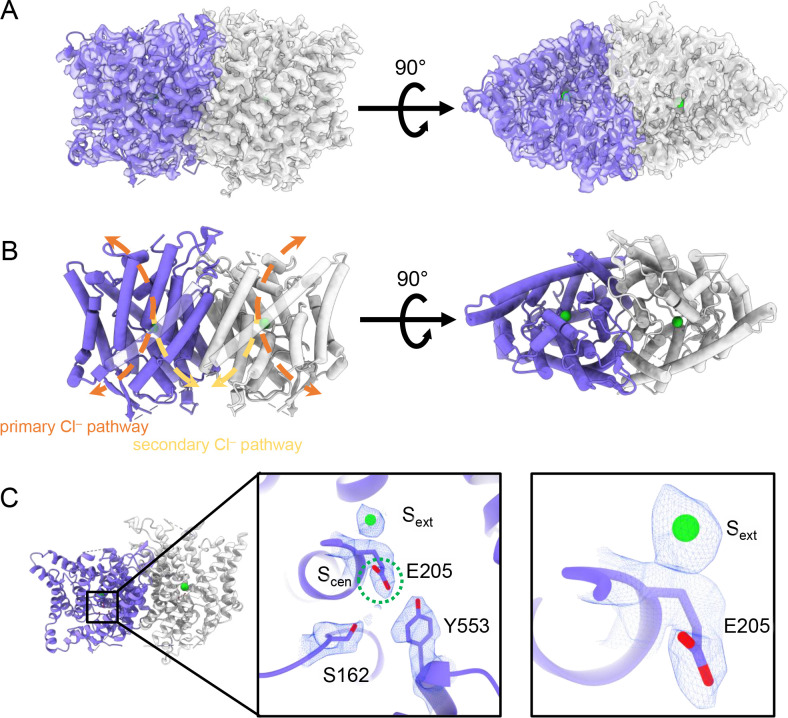
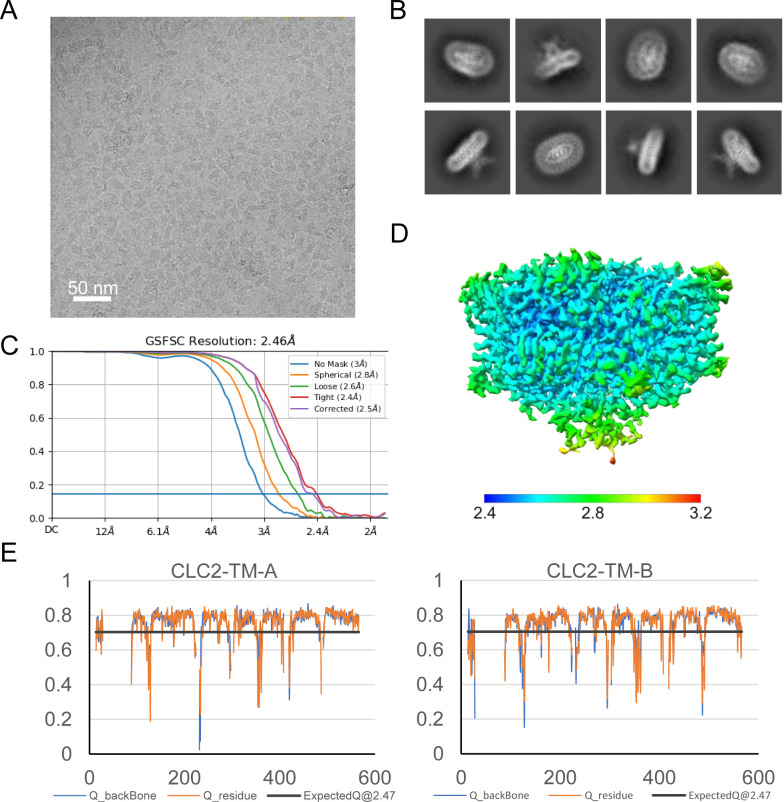
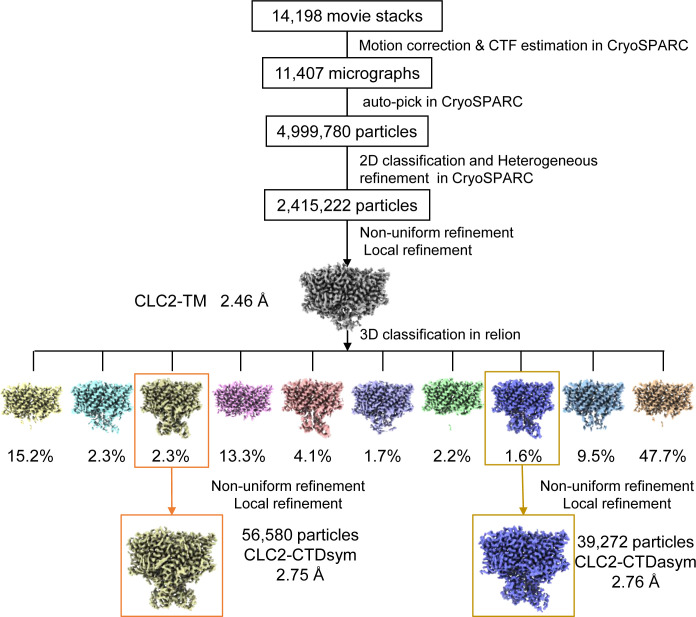
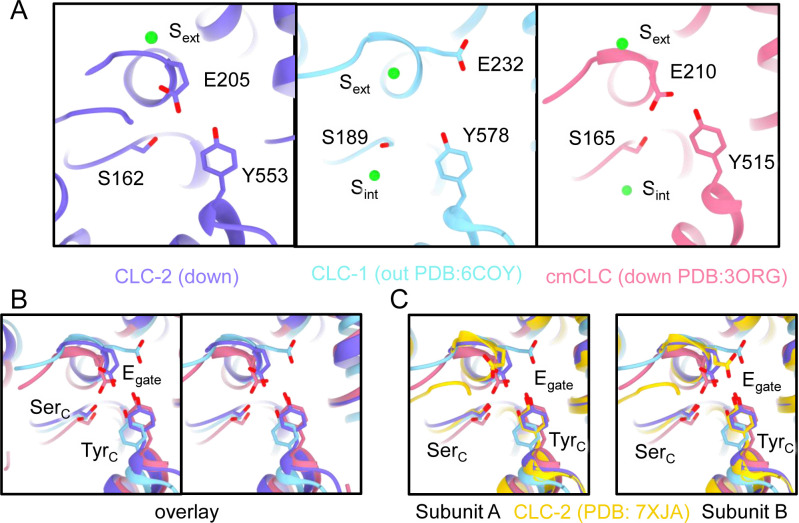
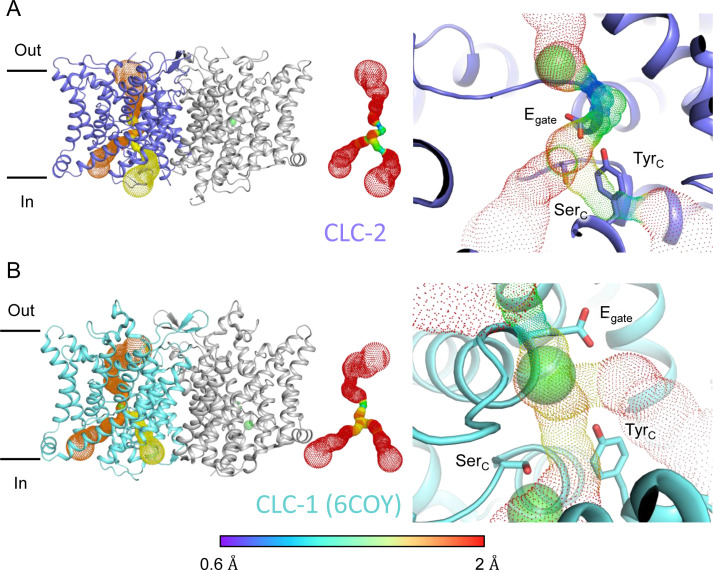
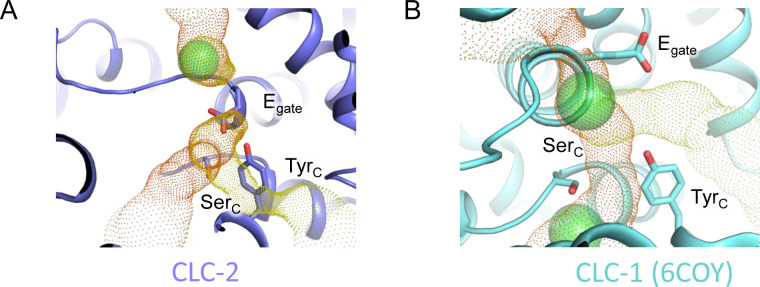
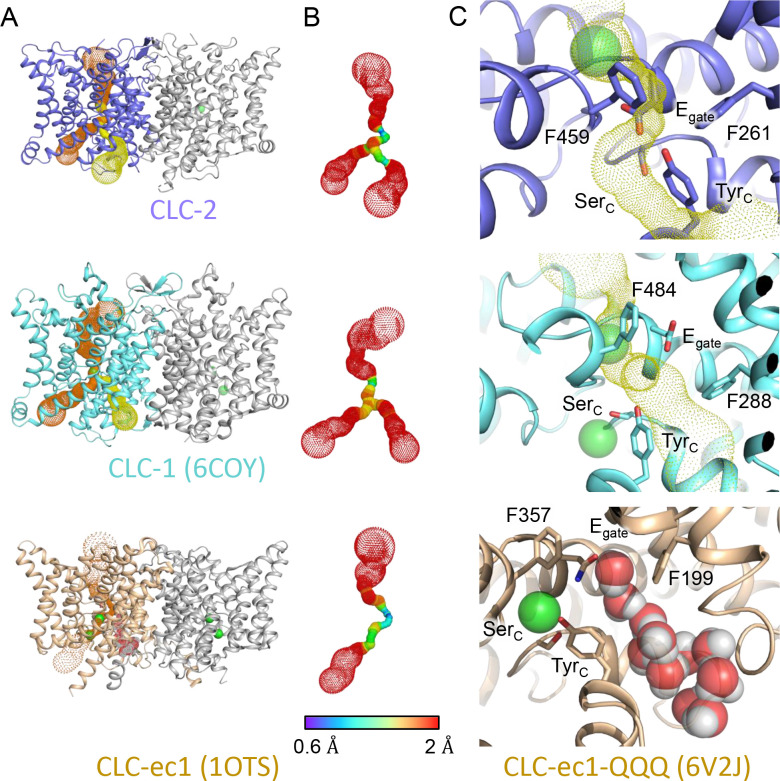
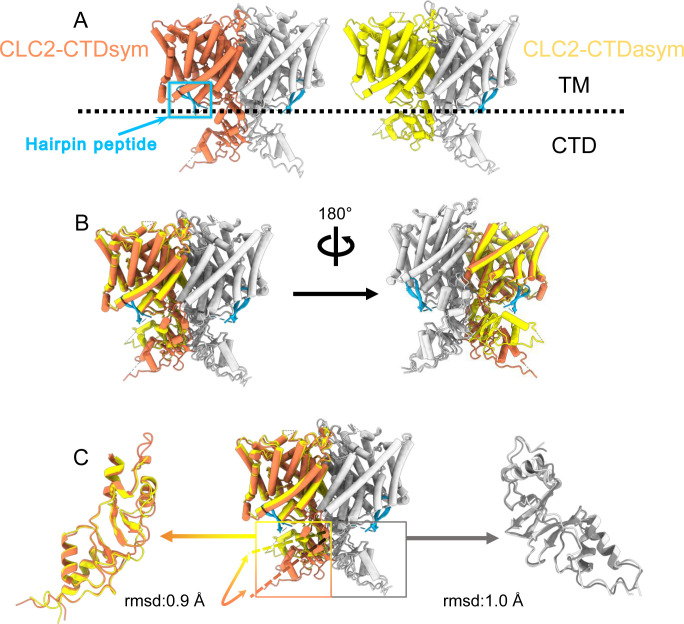
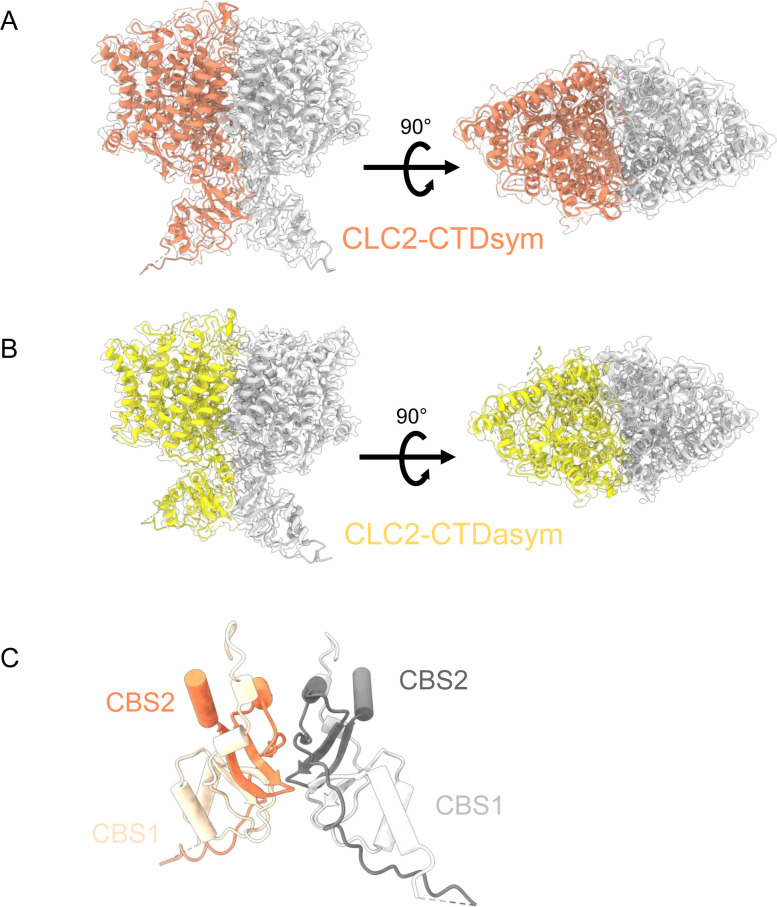
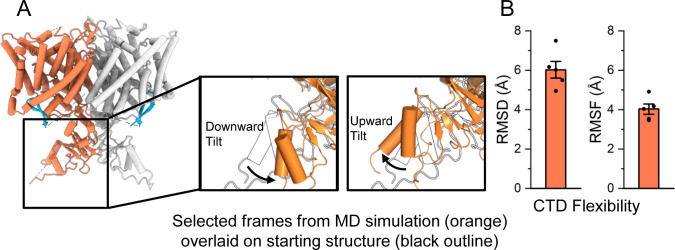
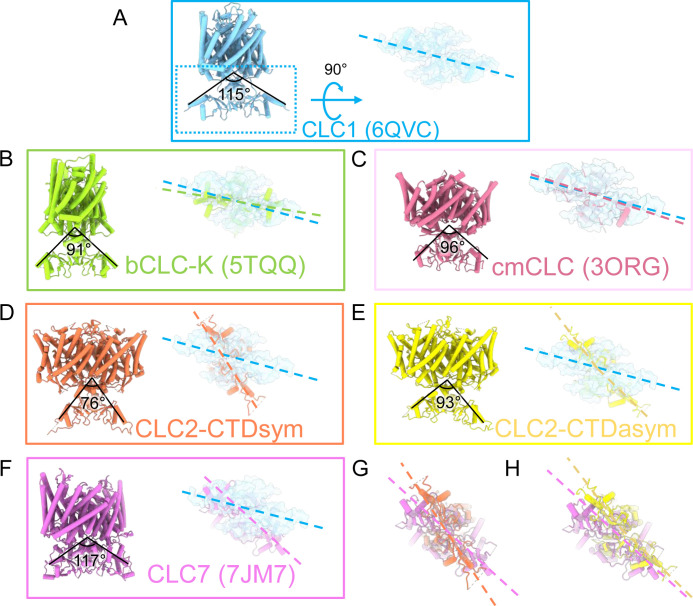
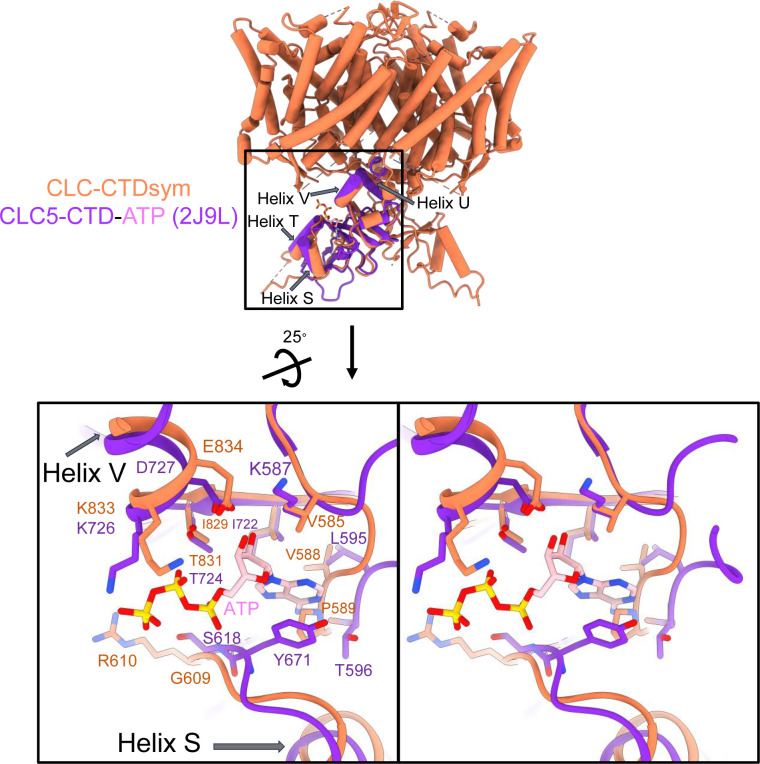
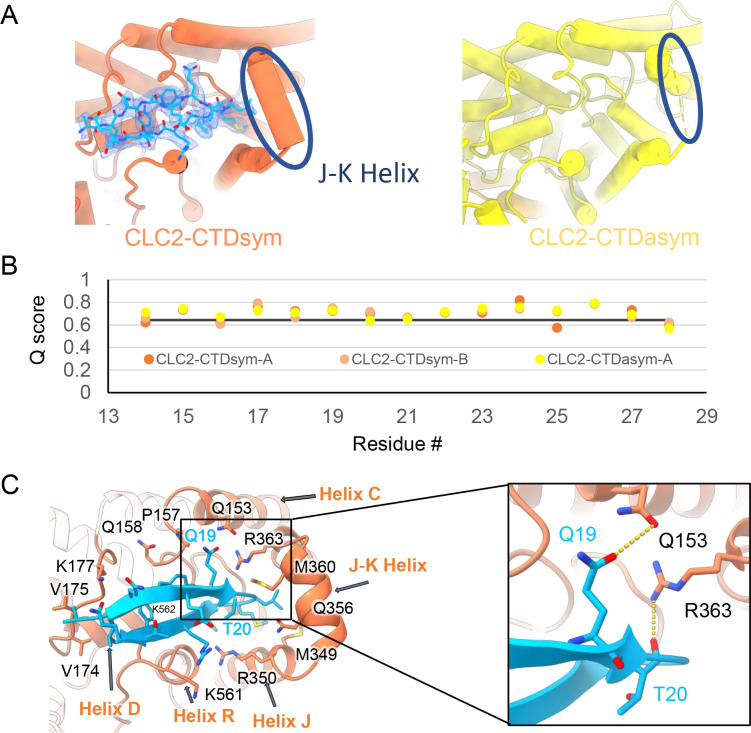
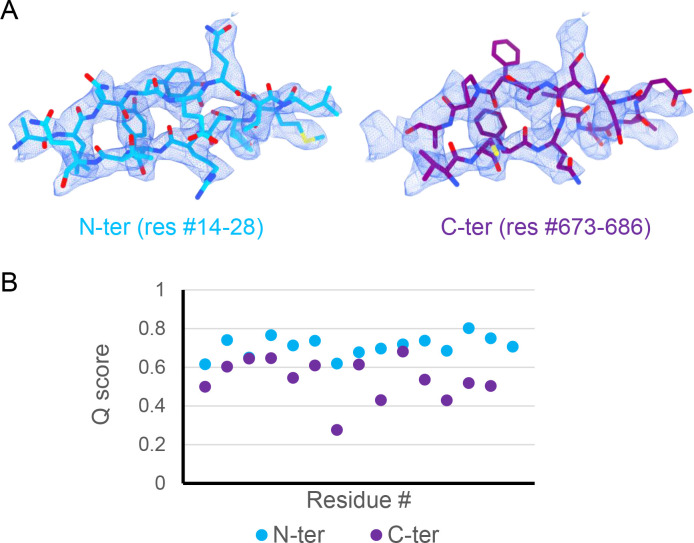
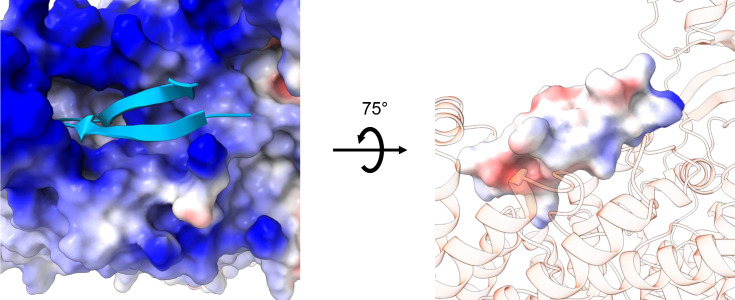
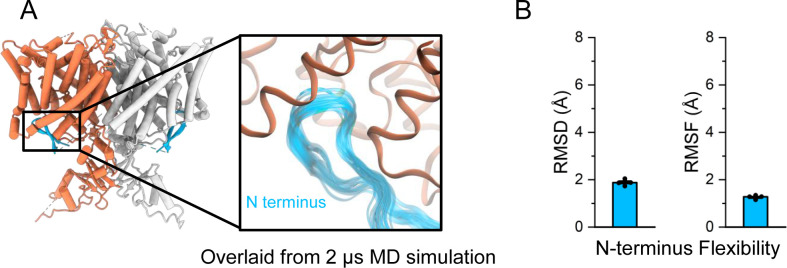
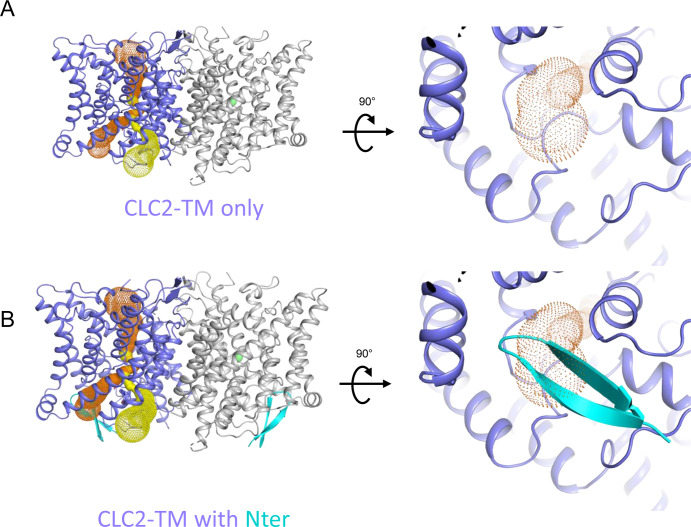
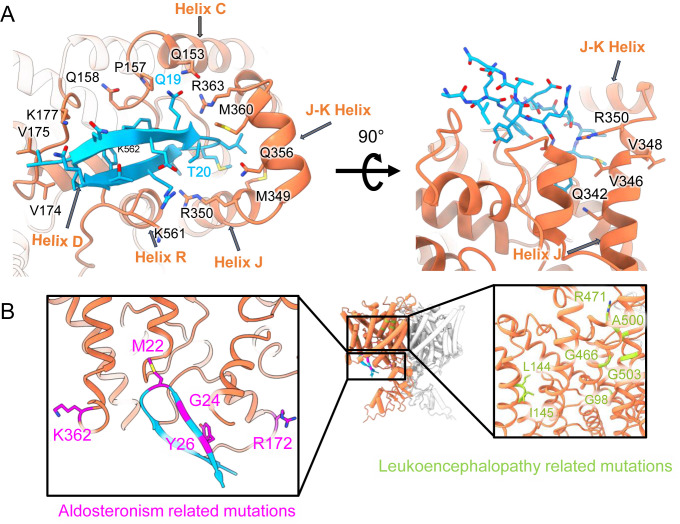
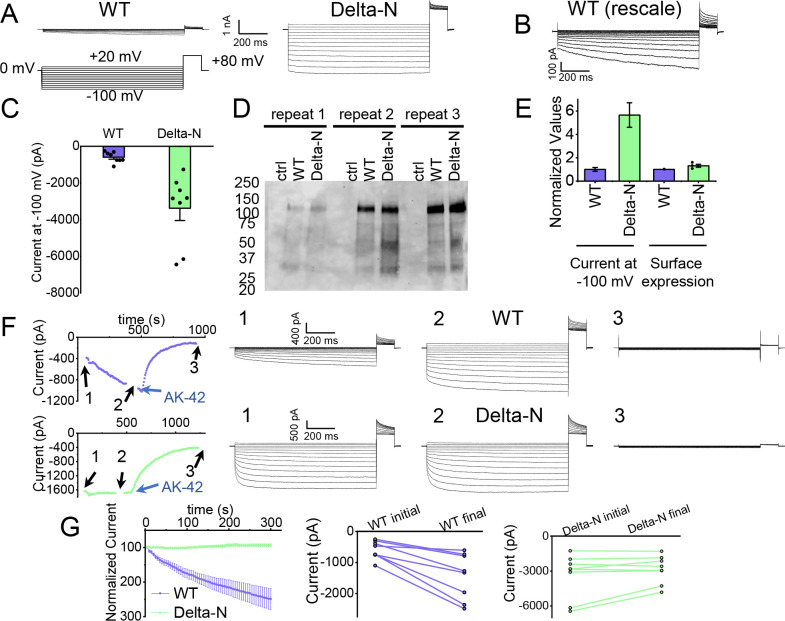
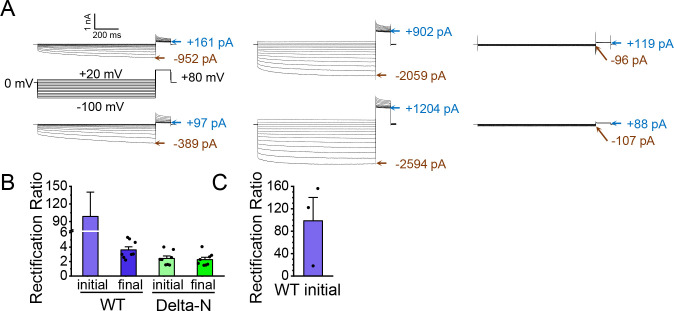
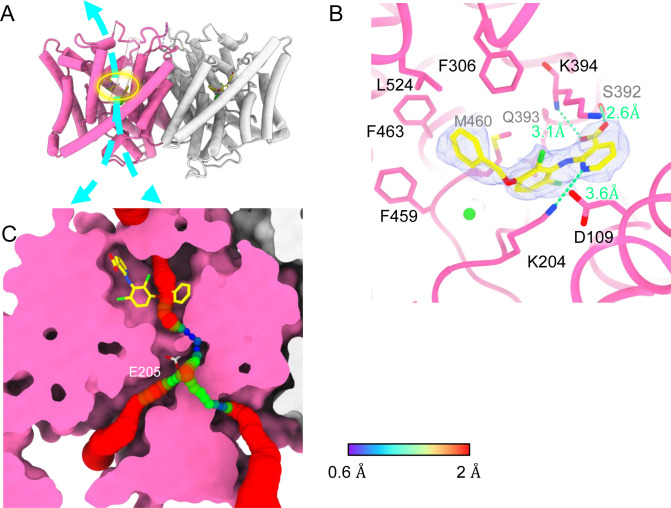
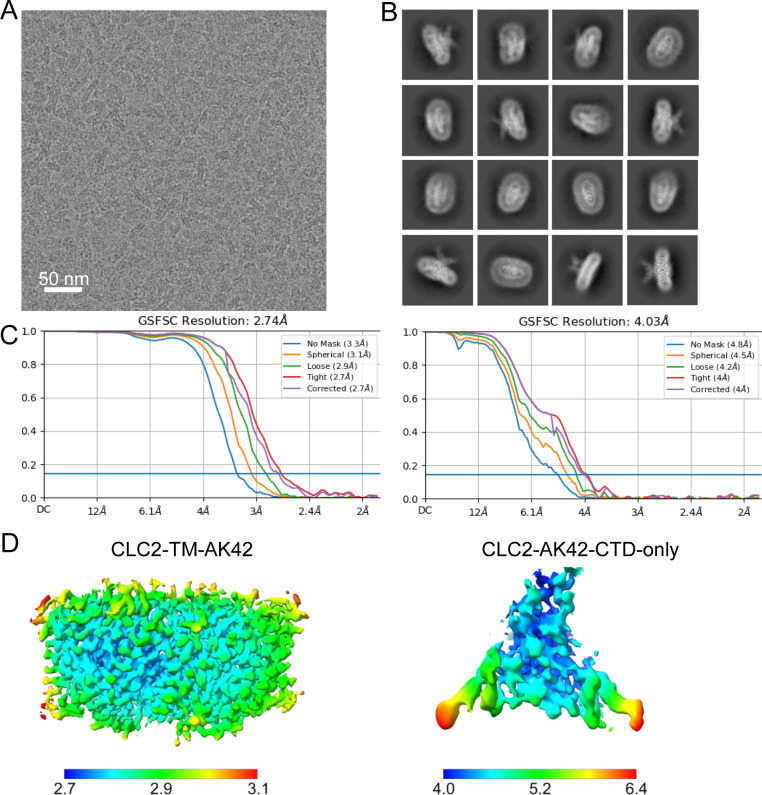
CLC-2 is a voltage-gated chloride channel that contributes to electrical excitability and ion homeostasis in many different tissues. Among the nine mammalian CLC homologs, CLC-2 is uniquely activated by hyperpolarization, rather than depolarization, of the plasma membrane. The molecular basis for the divergence in polarity of voltage gating among closely related homologs has been a long-standing mystery, in part because few CLC channel structures are available. Here, we report cryoEM structures of human CLC-2 at 2.46 - 2.76 Å, in the presence and absence of the selective inhibitor AK-42. AK-42 binds within the extracellular entryway of the Cl-permeation pathway, occupying a pocket previously proposed through computational docking studies. In the apo structure, we observed two distinct conformations involving rotation of one of the cytoplasmic C-terminal domains (CTDs). In the absence of CTD rotation, an intracellular N-terminal 15-residue hairpin peptide nestles against the TM domain to physically occlude the Cl-permeation pathway. This peptide is highly conserved among species variants of CLC-2 but is not present in other CLC homologs. Previous studies suggested that the N-terminal domain of CLC-2 influences channel properties via a "ball-and-chain" gating mechanism, but conflicting data cast doubt on such a mechanism, and thus the structure of the N-terminal domain and its interaction with the channel has been uncertain. Through electrophysiological studies of an N-terminal deletion mutant lacking the 15-residue hairpin peptide, we support a model in which the N-terminal hairpin of CLC-2 stabilizes a closed state of the channel by blocking the cytoplasmic Cl-permeation pathway.
CLC-2 是一种电压门控氯离子通道,它有助于许多不同组织的电兴奋性和离子内稳态。在哺乳动物的 9 种 CLC 同源物中,CLC-2 独特地被细胞膜的超极化而不是去极化激活。在密切相关的同源物中,电压门控极性差异的分子基础一直是一个长期存在的谜,部分原因是很少有 CLC 通道结构可用。在这里,我们报告了在存在和不存在选择性抑制剂 AK-42 的情况下,人 CLC-2 的 2.46-2.76Å 的冷冻电镜结构。AK-42 结合在氯离子渗透途径的细胞外入口内,占据了以前通过计算对接研究提出的口袋。在apo 结构中,我们观察到两种涉及一个细胞质 C 端结构域(CTD)旋转的不同构象。在没有 CTD 旋转的情况下,一个细胞内的 N 端 15 个残基发夹肽紧贴 TM 域,物理上阻塞了氯离子渗透途径。该肽在 CLC-2 的物种变体中高度保守,但在其他 CLC 同源物中不存在。以前的研究表明,CLC-2 的 N 端结构域通过“球链”门控机制影响通道特性,但相互矛盾的数据对这种机制提出了质疑,因此 N 端结构域及其与通道的相互作用一直不确定。通过对缺失了 15 个残基发夹肽的 N 端缺失突变体的电生理研究,我们支持这样一种模型,即 CLC-2 的 N 端发夹通过阻塞细胞质氯离子渗透途径来稳定通道的关闭状态。