Hansel Christian, Yuste Rafael
Department of Neurobiology and Neuroscience Institute, University of Chicago, Chicago, IL, United States.
NeuroTechnology Center, Department of Biological Sciences, Columbia University, New York, NY, United States.
Front Cell Neurosci. 2024 Jul 31;18:1440588. doi: 10.3389/fncel.2024.1440588. eCollection 2024.
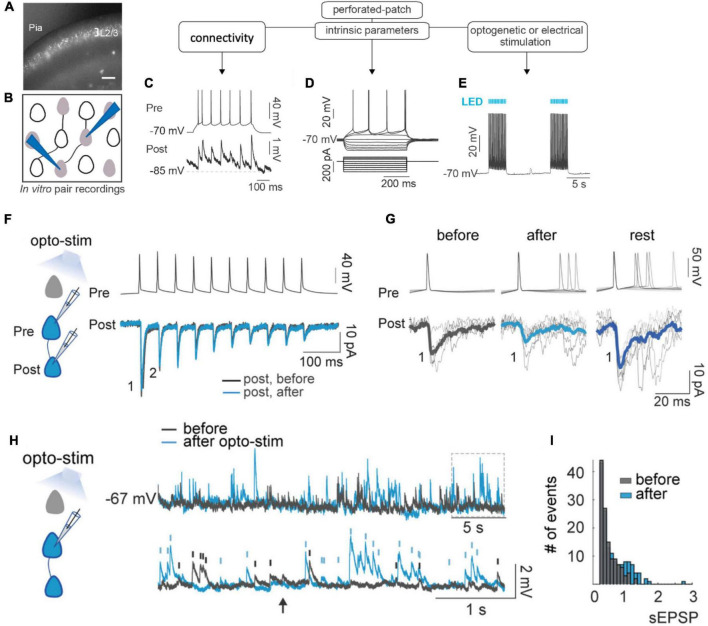
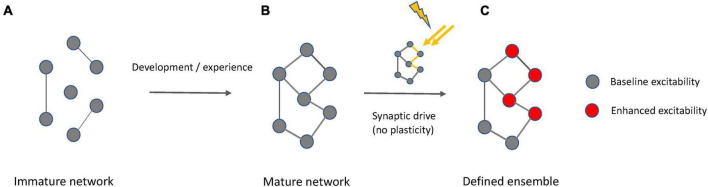
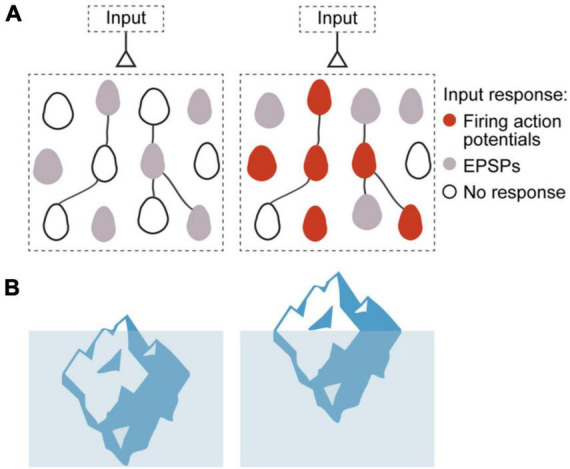
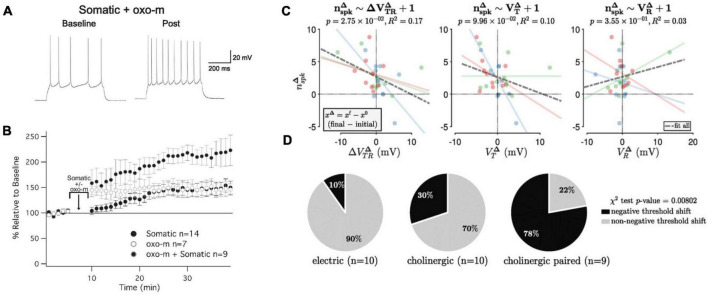
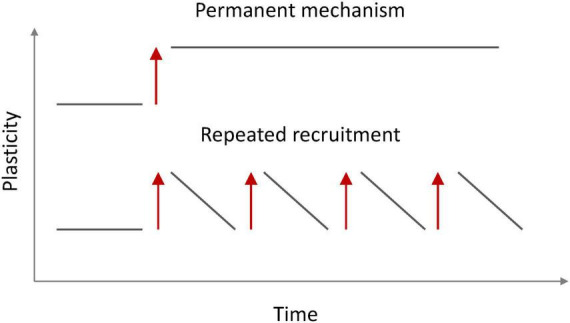
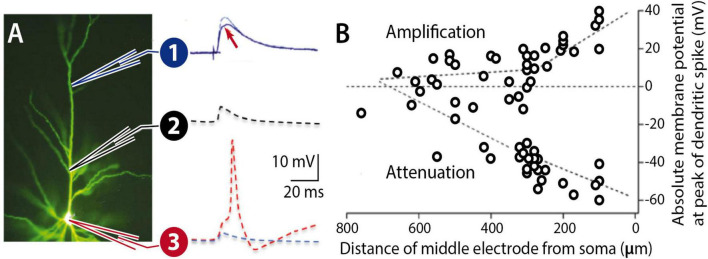
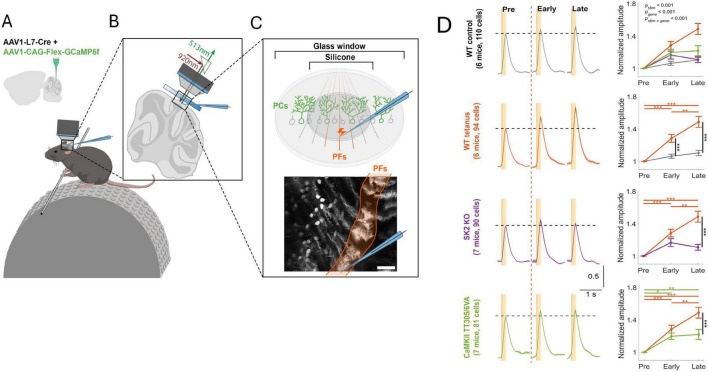
Synaptic connectivity defines groups of neurons that engage in correlated activity during specific functional tasks. These co-active groups of neurons form ensembles, the operational units involved in, for example, sensory perception, motor coordination and memory (then called an ). Traditionally, ensemble formation has been thought to occur via strengthening of synaptic connections via long-term potentiation (LTP) as a plasticity mechanism. This synaptic theory of memory arises from the learning rules formulated by Hebb and is consistent with many experimental observations. Here, we propose, as an alternative, that the intrinsic excitability of neurons and its plasticity constitute a second, mechanism that could be important for the initial formation of ensembles. Indeed, enhanced neural excitability is widely observed in multiple brain areas subsequent to behavioral learning. In cortical structures and the amygdala, excitability changes are often reported as transient, even though they can last tens of minutes to a few days. Perhaps it is for this reason that they have been traditionally considered as modulatory, merely supporting ensemble formation by facilitating LTP induction, without further involvement in memory function (memory allocation hypothesis). We here suggest-based on two lines of evidence-that beyond modulating LTP allocation, enhanced excitability plays a more fundamental role in learning. First, enhanced excitability constitutes a signature of active ensembles and, due to it, subthreshold synaptic connections become suprathreshold in the absence of synaptic plasticity (). Second, enhanced excitability promotes the propagation of dendritic potentials toward the soma and allows for enhanced coupling of EPSP amplitude (LTP) to the spike output (and thus ensemble participation). This describes a need for permanently increased excitability, which seems at odds with its traditional consideration as a short-lived mechanism. We propose that longer modifications in excitability are made possible by a low threshold for intrinsic plasticity induction, suggesting that excitability might be on/off-modulated at short intervals. Consistent with this, in cerebellar Purkinje cells, excitability lasts days to weeks, which shows that in some circuits the duration of the phenomenon is not a limiting factor in the first place. In our model, synaptic plasticity defines the information content received by neurons through the connectivity network that they are embedded in. However, the plasticity of cell-autonomous excitability could dynamically regulate the ensemble participation of individual neurons as well as the overall activity state of an ensemble.
突触连接性定义了在特定功能任务中参与相关活动的神经元群体。这些共同激活的神经元群体形成了集合,即参与例如感觉感知、运动协调和记忆(此时称为 )的操作单元。传统上,人们认为集合的形成是通过作为可塑性机制的长时程增强(LTP)来加强突触连接实现的。这种记忆的突触理论源于赫布提出的学习规则,并且与许多实验观察结果一致。在这里,我们提出另一种观点,即神经元的内在兴奋性及其可塑性构成了第二个 机制,这可能对集合的初始形成很重要。事实上,在行为学习后,多个脑区广泛观察到神经兴奋性增强。在皮质结构和杏仁核中,兴奋性变化通常被报道为短暂的,尽管它们可以持续数十分钟到几天。也许正是因为这个原因,它们传统上被认为是调节性的,仅仅通过促进LTP诱导来支持集合形成,而不进一步参与记忆功能(记忆分配假说)。基于两条证据,我们在此表明,除了调节LTP分配外,增强的兴奋性在学习中发挥着更基本的作用。首先,增强的兴奋性构成了活跃集合的特征,并且由于它,在没有突触可塑性的情况下,阈下突触连接会变为阈上连接( )。其次,增强的兴奋性促进树突电位向胞体的传播,并允许增强兴奋性突触后电位(EPSP)幅度(LTP)与动作电位输出(从而参与集合)的耦合。这 描述了对永久性增加兴奋性的需求,这似乎与它传统上被视为短暂机制的观点相矛盾。我们提出,通过内在可塑性诱导的低阈值,可以实现兴奋性的更长时间改变,这表明兴奋性可能在短时间间隔内进行开/关调节。与此一致的是,在小脑浦肯野细胞中,兴奋性持续数天到数周,这表明在某些回路中,该现象的持续时间首先不是一个限制因素。在我们的模型中,突触可塑性通过神经元所嵌入的连接网络定义了神经元接收到的信息内容。然而,细胞自主兴奋性的可塑性可以动态调节单个神经元的集合参与以及集合的整体活动状态。