Wang J, Smerdon S J, Jäger J, Kohlstaedt L A, Rice P A, Friedman J M, Steitz T A
Department of Molecular Biophysics and Biochemistry, Howard Hughes Medical Institute, Yale University New Haven, CT 06520-8114.
Proc Natl Acad Sci U S A. 1994 Jul 19;91(15):7242-6. doi: 10.1073/pnas.91.15.7242.
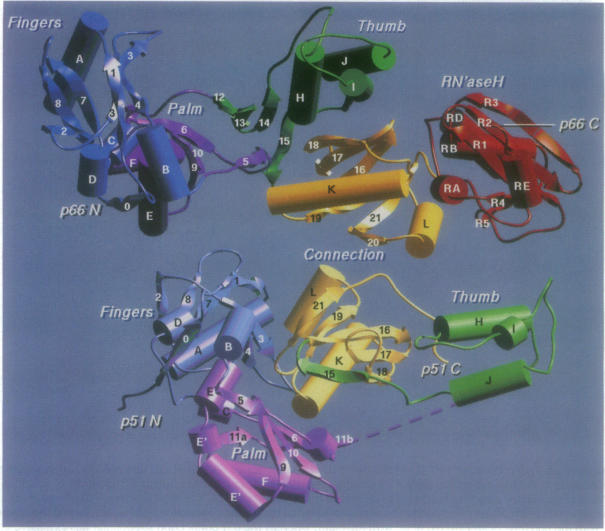
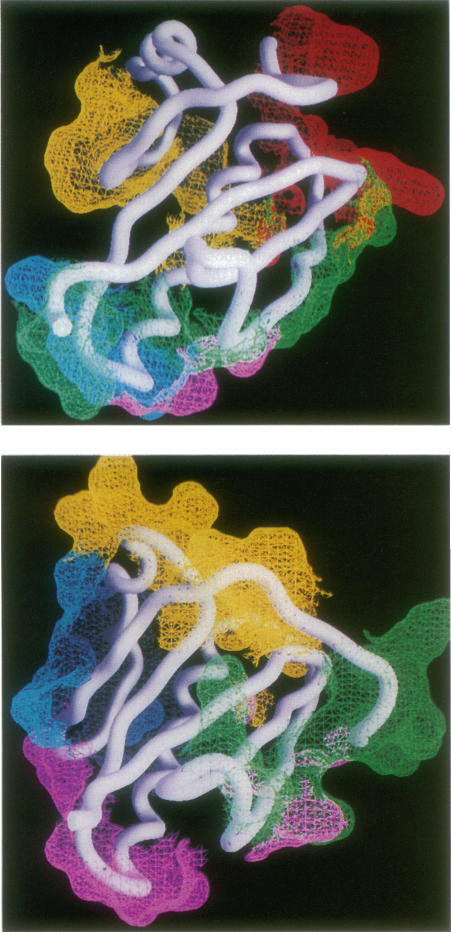
The reverse transcriptase from human immunodeficiency virus type 1 is a heterodimer consisting of one 66-kDa and one 51-kDa subunit. The p66 subunit contains both a polymerase and an RNase H domain; proteolytic cleavage of p66 removes the RNase H domain to yield the p51 subunit. Although the polymerase domain of p66 folds into an open, extended structure containing a large active-site cleft, that of p51 is closed and compact. The connection subdomain, which lies between the polymerase and RNase H active sites in p66, plays a central role in the formation of the reverse transcriptase heterodimer. Extensive and very different intra- and intersubunit contacts are made by the connection subdomains of each of the subunits. Together, contacts between the two connection domains constitute approximately one-third of the total contacts between subunits of the heterodimer. Conversion of an open p66 polymerase domain structure to a closed p51-like structure results in a reduction in solvent-accessible surface area by 1600 A2 and the burying of an extensive hydrophobic surface. Thus, the monomeric forms of both p66 and p51 are proposed to have the same closed structure as seen in the p51 subunit of the heterodimer. The free energy required to convert p66 from a closed p51-like structure to the observed open p66 polymerase domain structure is generated by the burying of a large, predominantly hydrophobic surface area upon formation of the heterodimer. It is likely that the only kind of dimer that can form is an asymmetric one like that seen in the heterodimer structure, since one dimer interaction surface exists only in p51 and the other only in p66. We suggest that both p51 and p66 form asymmetric homodimers that are assembled from one subunit that has assumed the open conformation and one that has the closed structure.
1型人类免疫缺陷病毒的逆转录酶是一种异源二聚体,由一个66 kDa亚基和一个51 kDa亚基组成。p66亚基同时包含一个聚合酶结构域和一个核糖核酸酶H结构域;p66经蛋白水解切割后去除核糖核酸酶H结构域,产生p51亚基。尽管p66的聚合酶结构域折叠成一个开放的、伸展的结构,包含一个大的活性位点裂缝,但p51的聚合酶结构域是封闭且紧凑的。连接亚结构域位于p66的聚合酶和核糖核酸酶H活性位点之间,在逆转录酶异源二聚体的形成中起核心作用。每个亚基的连接亚结构域形成了广泛且非常不同的亚基内和亚基间接触。两个连接结构域之间的接触共同构成了异源二聚体亚基间总接触的约三分之一。将开放的p66聚合酶结构域结构转变为封闭的p51样结构会导致溶剂可及表面积减少1600 Ų,并掩埋一个广泛的疏水表面。因此,有人提出p66和p51的单体形式具有与异源二聚体p51亚基中所见相同的封闭结构。将p66从封闭的p51样结构转变为观察到的开放p66聚合酶结构域结构所需的自由能是由异源二聚体形成时掩埋一个大的、主要为疏水的表面积产生的。可能形成的唯一一种二聚体是像在异源二聚体结构中看到的不对称二聚体,因为一个二聚体相互作用表面仅存在于p51中,另一个仅存在于p66中。我们认为p51和p66都形成不对称同源二聚体,这些二聚体由一个呈开放构象的亚基和一个具有封闭结构的亚基组装而成。