Kim E, Du L, Bregman D B, Warren S L
Department of Pathology, Yale University School of Medicine, New Haven, Connecticut 06510, USA.
J Cell Biol. 1997 Jan 13;136(1):19-28. doi: 10.1083/jcb.136.1.19.
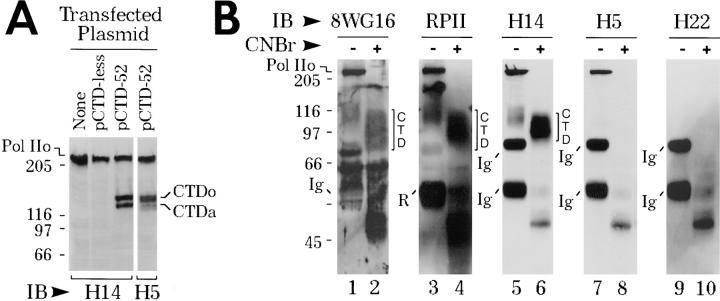
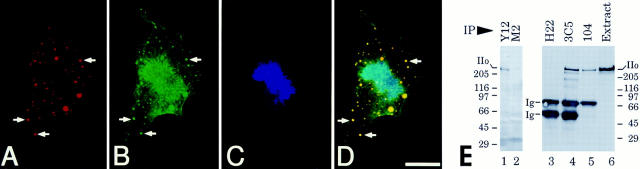
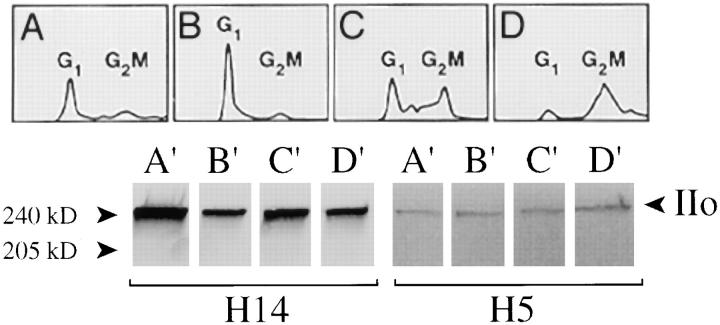
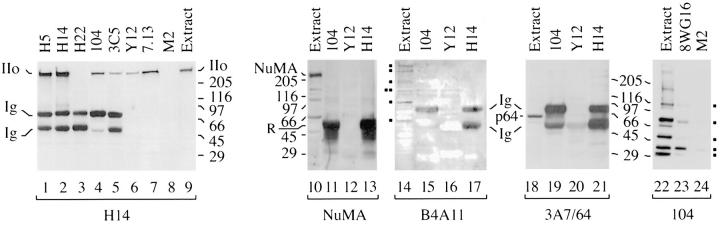
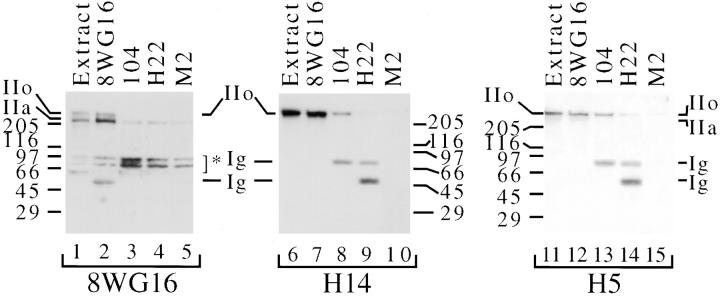
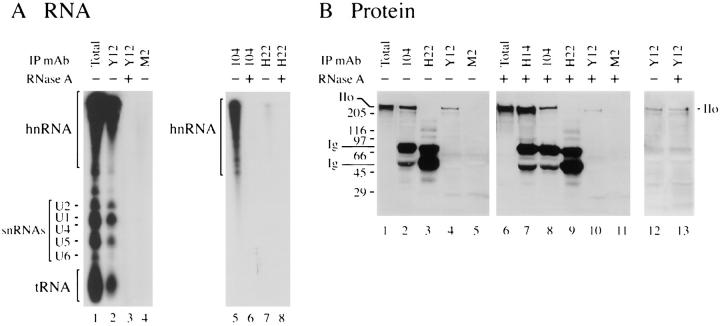
The carboxy-terminal domain (CTD) of the largest subunit of RNA polymerase II (Pol II) contains multiple tandem copies of the consensus heptapeptide, TyrSerProThrSerProSer. Concomitant with transcription initiation the CTD is phosphorylated. Elongating polymerase has a hyperphosphorylated CTD, but the role of this modification is poorly understood. A recent study revealed that some hyperphosphorylated polymerase molecules (Pol IIo) are nonchromosomal, and hence transcriptionally unengaged (Bregman, D.B., L. Du, S. van der Zee, S.L. Warren. 1995. J. Cell Biol. 129: 287-298). Pol IIo was concentrated in discrete splicing factor domains, suggesting a possible relationship between CTD phosphorylation and splicing factors, but no evidence beyond immunolocalization data was provided to support this idea. Here, we show that Pol IIo co-immunoprecipitates with members of two classes of splicing factors, the Sm snRNPs and non-snRNP SerArg (SR) family proteins. Significantly, Pol IIo's association with splicing factors is maintained in the absence of pre-mRNA, and the polymerase need not be transcriptionally engaged. We also provide definitive evidence that hyperphosphorylation of Pol II's CTD is poorly correlated with its transcriptional activity. Using monoclonal antibodies (mAbs) H5 and H14, which are shown here to recognize phosphoepitopes on Pol II's CTD, we have quantitated the level of Pol IIo at different stages of the cell cycle. The level of Pol IIo is similar in interphase and mitotic cells, which are transcriptionally active and inactive, respectively. Finally, complexes containing Pol IIo and splicing factors can be prepared from mitotic as well as interphase cells. The experiments reported here establish that hyperphosphorylation of the CTD is a good indicator of polymerase's association with snRNP and SR splicing factors, but not of its transcriptional activity. Most importantly, the present study suggests that splicing factors may associate with the polymerase via the hyperphosphorylated CTD.
RNA聚合酶II(Pol II)最大亚基的羧基末端结构域(CTD)包含多个七肽共有序列TyrSerProThrSerProSer的串联重复。伴随着转录起始,CTD被磷酸化。延伸中的聚合酶具有高度磷酸化的CTD,但这种修饰的作用却知之甚少。最近的一项研究表明,一些高度磷酸化的聚合酶分子(Pol IIo)是非染色体的,因此没有参与转录(Bregman,D.B.,L. Du,S. van der Zee,S.L. Warren. 1995. 《细胞生物学杂志》129:287 - 298)。Pol IIo集中在离散的剪接因子结构域中,这表明CTD磷酸化与剪接因子之间可能存在某种关系,但除了免疫定位数据外,没有提供其他证据来支持这一观点。在此,我们发现Pol IIo能与两类剪接因子的成员共免疫沉淀,即Sm snRNP和非snRNP SerArg(SR)家族蛋白。值得注意的是,在没有前体mRNA的情况下,Pol IIo与剪接因子的结合依然存在,并且聚合酶无需参与转录。我们还提供了确凿的证据,表明Pol II的CTD高度磷酸化与其转录活性之间的相关性很差。使用单克隆抗体(mAb)H5和H14(在此显示它们可识别Pol II的CTD上的磷酸表位),我们对细胞周期不同阶段的Pol IIo水平进行了定量。Pol IIo的水平在间期细胞和有丝分裂细胞中相似,而这两类细胞分别处于转录活跃和不活跃状态。最后,含有Pol IIo和剪接因子的复合物可以从有丝分裂细胞以及间期细胞中制备出来。此处报道的实验证实,CTD的高度磷酸化是聚合酶与snRNP和SR剪接因子结合的良好指标,但并非其转录活性的指标。最重要的是,本研究表明剪接因子可能通过高度磷酸化的CTD与聚合酶结合。