Brierley I, Meredith M R, Bloys A J, Hagervall T G
Department of Pathology, University of Cambridge, UK.
J Mol Biol. 1997 Jul 18;270(3):360-73. doi: 10.1006/jmbi.1997.1134.
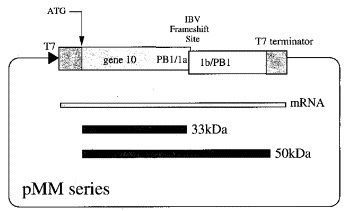
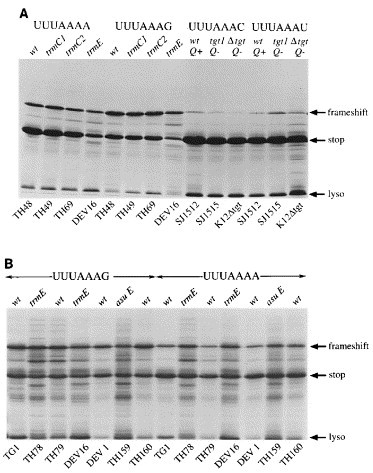
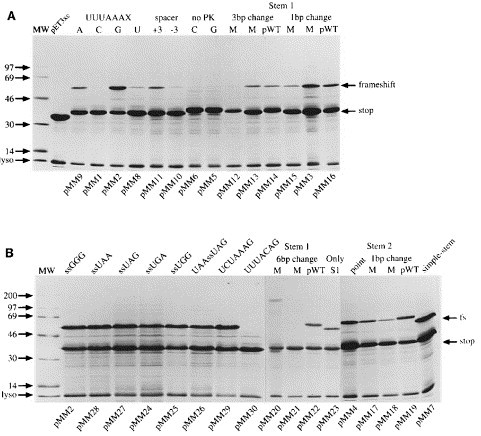
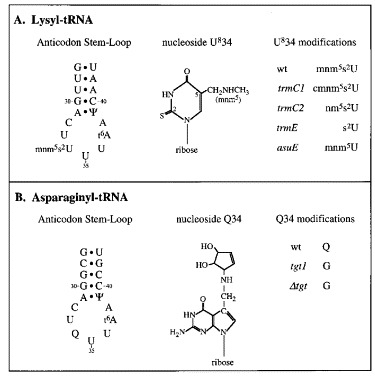
Eukaryotic ribosomal frameshift signals generally contain two elements, a heptanucleotide slippery sequence (XXXYYYN) and an RNA secondary structure, often an RNA pseudoknot, located downstream. Frameshifting takes place at the slippery sequence by simultaneous slippage of two ribosome-bound tRNAs. All of the tRNAs that are predicted to decode frameshift sites in the ribosomal A-site (XXXYYYN) possess a hypermodified base in the anticodon-loop and it is conceivable that these modifications play a role in the frameshift process. To test this, we expressed slippery sequence variants of the coronavirus IBV frameshift signal in strains of Escherichia coli unable to modify fully either tRNA(Lys) or tRNA(Asn). At the slippery sequences UUUAAAC and UUUAAAU (underlined codon decoded by tRNA(Asn), anticodon 5' QUU 3'), frameshifting was very inefficient (2 to 3%) and in strains deficient in the biosynthesis of Q base, was increased (AAU) or decreased (AAC) only two-fold. In E. coli, therefore, hypomodification of tRNA(Asn) had little effect on frameshifting. The situation with the efficient slippery sequences UUUAAAA (15%) and UUUAAAG (40%) (underlined codon decoded by tRNA(Lys), anticodon 5' mnm5s2UUU 3') was more complex, since the wobble base of tRNA(Lys) is modified at two positions. Of four available mutants, only trmE (s2UUU) had a marked influence on frameshifting, increasing the efficiency of the process at the slippery sequence UUUAAAA. No effect on frameshifting was seen in trmC1 (cmnm5s2UUU) or trmC2 (nm5s2UUU) strains and only a very small reduction (at UUUAAAG) was observed in an asuE (mnm5UUU) strain. The slipperiness of tRNA(Lys), therefore, cannot be ascribed to a single modification site on the base. However, the data support a role for the amino group of the mnm5 substitution in shaping the anticodon structure. Whether these conclusions can be extended to eukaryotic translation systems is uncertain. Although E. coli ribosomes changed frame at the IBV signal (UUUAAAG) with an efficiency similar to that measured in reticulocyte lysates (40%), there were important qualitative differences. Frameshifting of prokaryotic ribosomes was pseudoknot-independent (although secondary structure dependent) and appeared to require slippage of only a single tRNA.
真核生物核糖体移码信号通常包含两个元件,一个七核苷酸滑动序列(XXXYYYN)和一个位于下游的RNA二级结构,通常是一个RNA假结。移码发生在滑动序列处,由两个与核糖体结合的tRNA同时滑动引起。所有预计在核糖体A位点(XXXYYYN)解码移码位点的tRNA在反密码子环中都有一个高度修饰的碱基,可以想象这些修饰在移码过程中起作用。为了验证这一点,我们在无法完全修饰tRNA(Lys)或tRNA(Asn)的大肠杆菌菌株中表达了冠状病毒IBV移码信号的滑动序列变体。在滑动序列UUUAAAC和UUUAAAU(带下划线的密码子由tRNA(Asn)解码,反密码子5'QUU 3')处,移码效率非常低(2%至3%),在缺乏Q碱基生物合成的菌株中,仅增加(AAU)或降低(AAC)两倍。因此,在大肠杆菌中,tRNA(Asn)的低修饰对移码影响很小。高效滑动序列UUUAAAA(15%)和UUUAAAG(40%)(带下划线的密码子由tRNA(Lys)解码,反密码子5'mnm5s2UUU 3')的情况更为复杂,因为tRNA(Lys)的摆动碱基在两个位置被修饰。在四个可用的突变体中,只有trmE(s2UUU)对移码有显著影响,提高了在滑动序列UUUAAAA处的移码效率。在trmC1(cmnm5s2UUU)或trmC2(nm5s2UUU)菌株中未观察到对移码的影响,在asuE(mnm5UUU)菌株中仅观察到非常小的降低(在UUUAAAG处)。因此,tRNA(Lys)的滑动性不能归因于碱基上的单个修饰位点。然而,数据支持mnm5取代的氨基在塑造反密码子结构中的作用。这些结论是否可以扩展到真核生物翻译系统尚不确定。尽管大肠杆菌核糖体在IBV信号(UUUAAAG)处改变读框的效率与在网织红细胞裂解物中测得的效率相似(40%),但存在重要的质的差异。原核生物核糖体的移码与假结无关(尽管依赖二级结构),并且似乎只需要单个tRNA的滑动。