Bloch W, Forsberg E, Lentini S, Brakebusch C, Martin K, Krell H W, Weidle U H, Addicks K, Fässler R
Institute for Anatomy, University of Cologne, 50931 Cologne, Germany.
J Cell Biol. 1997 Oct 6;139(1):265-78. doi: 10.1083/jcb.139.1.265.
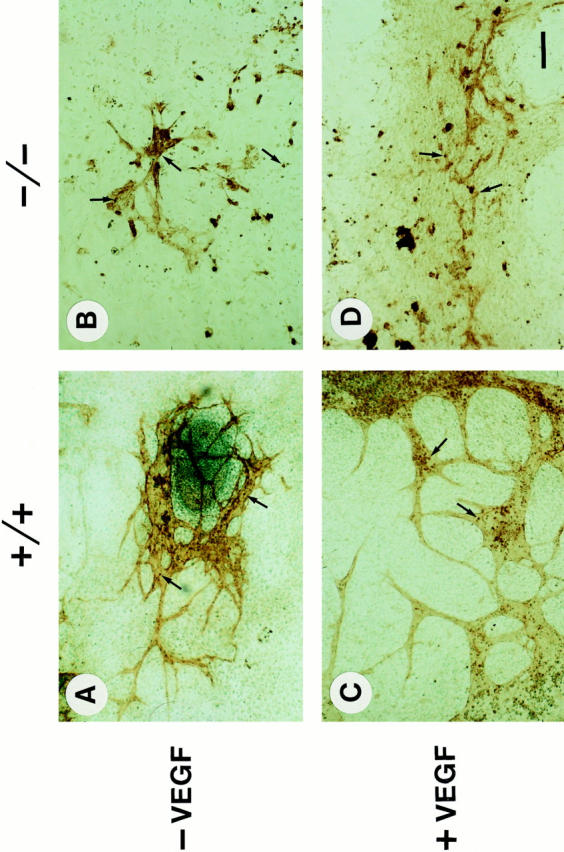
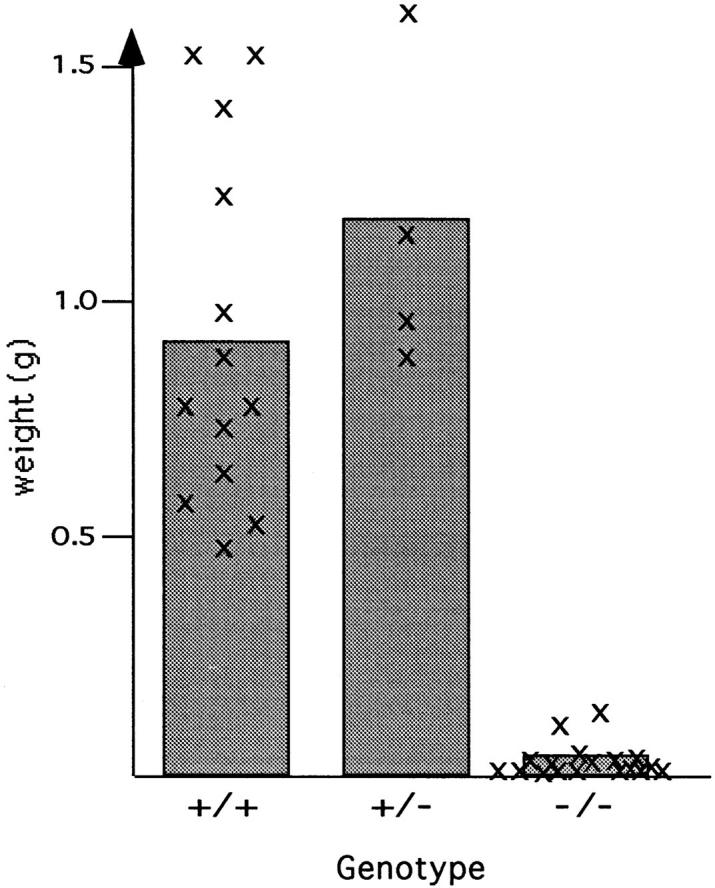
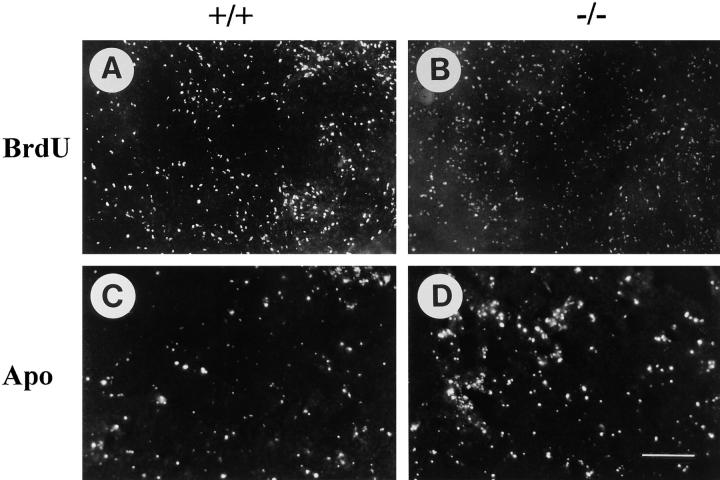
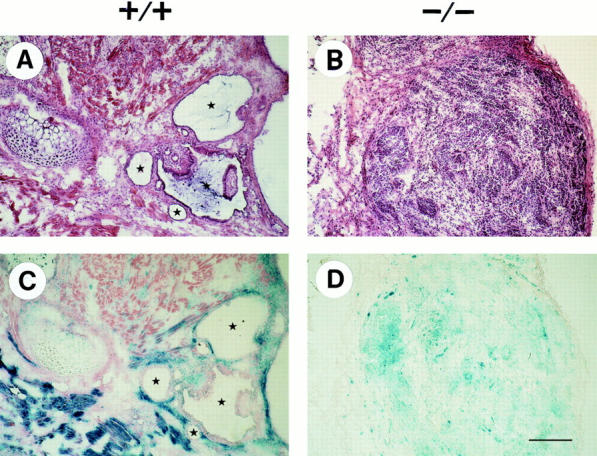
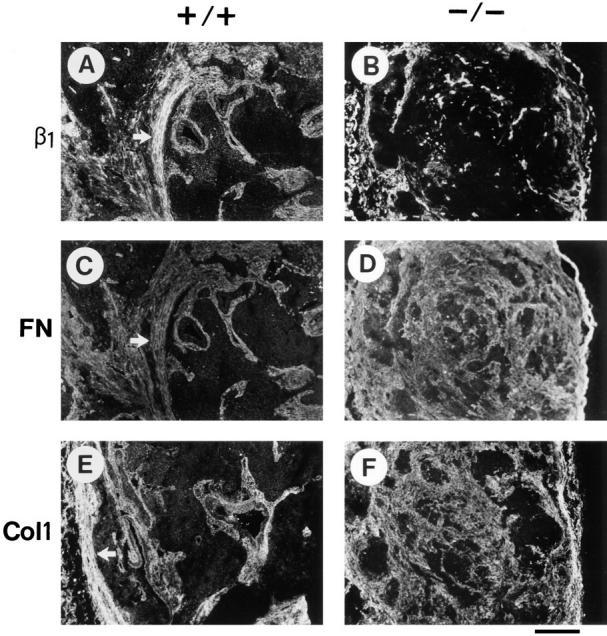
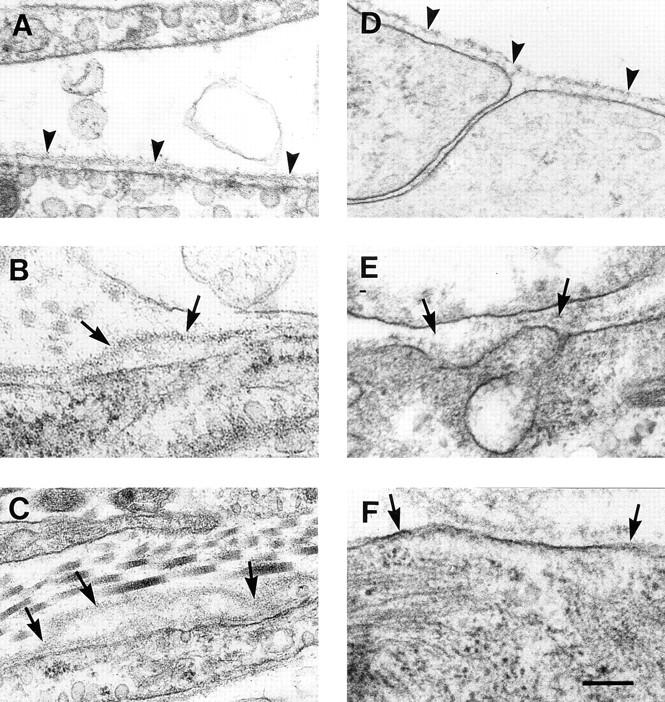
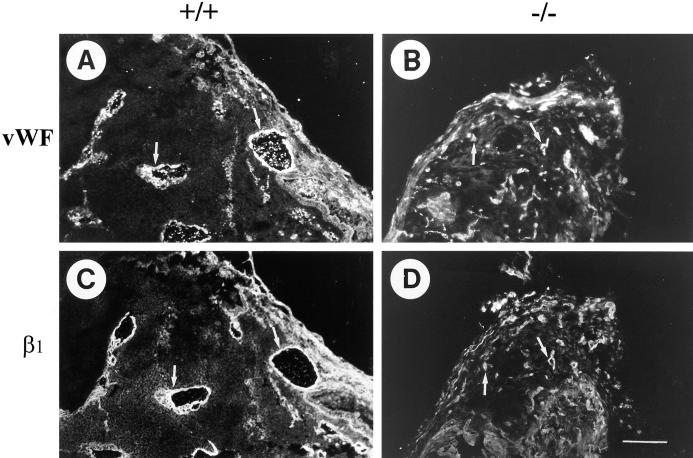
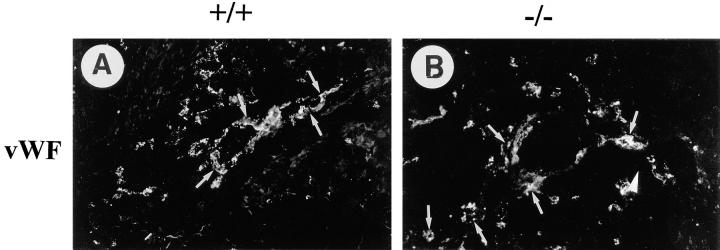
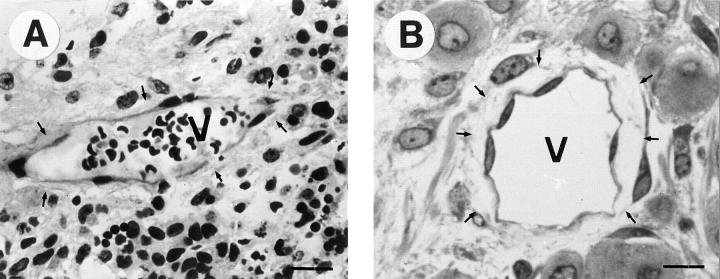
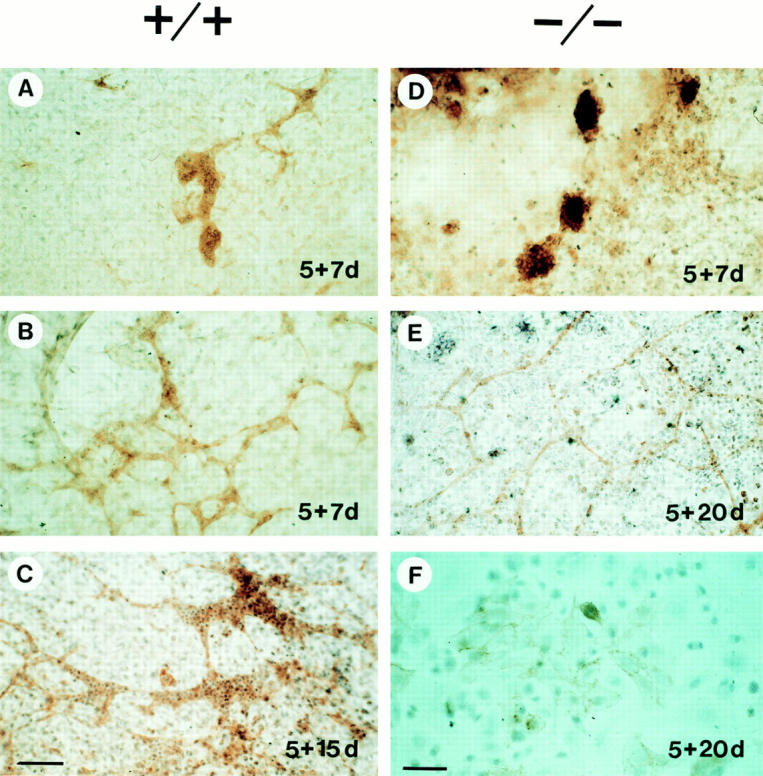
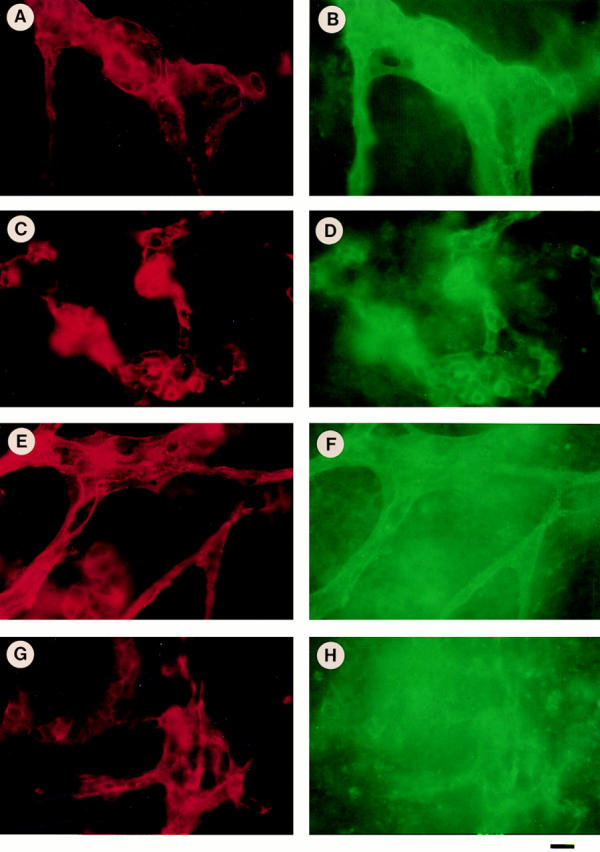
Teratomas are benign tumors that form after ectopic injection of embryonic stem (ES) cells into mice and contain derivatives of all primitive germ layers. To study the role of beta 1 integrin during teratoma formation, we compared teratomas induced by normal and beta1-null ES cells. Injection of normal ES cells gave rise to large teratomas. In contrast, beta 1-null ES cells either did not grow or formed small teratomas with an average weight of <5% of that of normal teratomas. Histological analysis of beta 1-null teratomas revealed the presence of various differentiated cells, however, a much lower number of host-derived stromal cells than in normal teratomas. Fibronectin, collagen I, and nidogen were expressed but, in contrast to normal teratomas, diffusely deposited in beta1-null teratomas. Basement membranes were present but with irregular shape and detached from the cell surface. Normal teratomas had large blood vessels with a smooth inner surface, containing both host- and ES cell-derived endothelial cells. In contrast, beta 1-null teratomas had small vessels that were loosely embedded into the connective tissue. Furthermore, endothelial cells were always of host-derived origin and formed blood vessels with an irregular inner surface. Although beta 1- deficient endothelial cells were absent in teratomas, beta 1-null ES cells could differentiate in vitro into endothelial cells. The formation of a complex vasculature, however, was significantly delayed and of poor quality in beta1-null embryoid bodies. Moreover, while vascular endothelial growth factor induced proliferation of endothelial cells as well as an extensive branching of blood vessels in normal embryoid bodies, it had no effect in beta 1-null embryoid bodies.
畸胎瘤是在将胚胎干细胞(ES细胞)异位注射到小鼠体内后形成的良性肿瘤,包含所有原始胚层的衍生物。为了研究β1整合素在畸胎瘤形成过程中的作用,我们比较了正常ES细胞和β1基因敲除ES细胞诱导形成的畸胎瘤。注射正常ES细胞会产生大的畸胎瘤。相比之下,β1基因敲除的ES细胞要么不生长,要么形成小的畸胎瘤,其平均重量不到正常畸胎瘤的5%。对β1基因敲除畸胎瘤的组织学分析显示存在各种分化细胞,然而,与正常畸胎瘤相比,宿主来源的基质细胞数量要少得多。纤连蛋白、I型胶原和巢蛋白表达,但与正常畸胎瘤不同,它们在β1基因敲除的畸胎瘤中呈弥漫性沉积。基底膜存在,但形状不规则且与细胞表面分离。正常畸胎瘤有大血管,内表面光滑,包含宿主和ES细胞来源的内皮细胞。相比之下,β