Anderson B L, Boldogh I, Evangelista M, Boone C, Greene L A, Pon L A
Department of Pathology, Columbia University College of Physicians and Surgeons, New York 10032, USA.
J Cell Biol. 1998 Jun 15;141(6):1357-70. doi: 10.1083/jcb.141.6.1357.
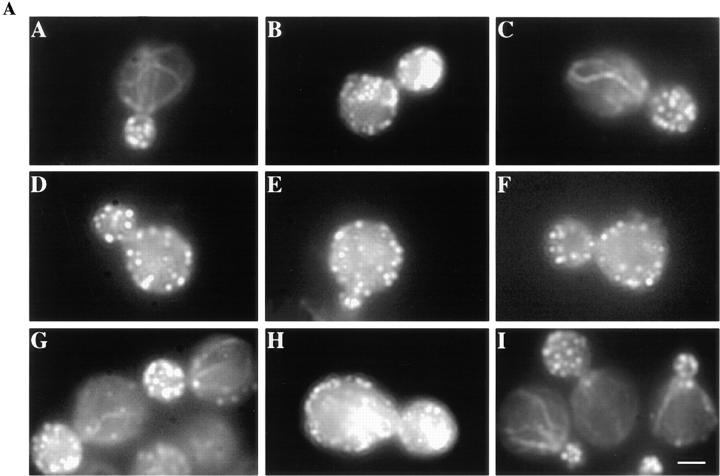
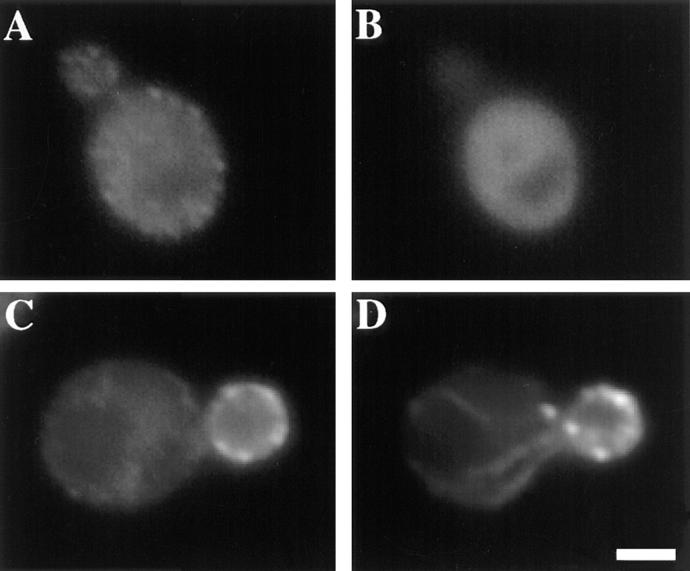
The budding yeast contains two type I myosins, Myo3p and Myo5p, with redundant functions. Deletion of both myosins results in growth defects, loss of actin polarity and polarized cell surface growth, and accumulation of intracellular membranes. Expression of myc-tagged Myo5p in myo3Delta myo5Delta cells fully restores wild-type characteristics. Myo5p is localized as punctate, cortical structures enriched at sites of polarized cell growth. We find that latrunculin-A-induced depolymerization of F-actin results in loss of Myo5p patches. Moreover, incubation of yeast cells at 37 degrees C results in transient depolarization of both Myo5p patches and the actin cytoskeleton. Mutant Myo5 proteins with deletions in nonmotor domains were expressed in myo3Delta myo5Delta cells and the resulting strains were analyzed for Myo5p function. Deletion of the tail homology 2 (TH2) domain, previously implicated in ATP-insensitive actin binding, has no detectable effect on Myo5p function. In contrast, myo3Delta myo5Delta cells expressing mutant Myo5 proteins with deletions of the src homology domain 3 (SH3) or both TH2 and SH3 domains display defects including Myo5p patch depolarization, actin disorganization, and phenotypes associated with actin dysfunction. These findings support a role for the SH3 domain in Myo5p localization and function in budding yeast. The proline-rich protein verprolin (Vrp1p) binds to the SH3 domain of Myo3p or Myo5p in two-hybrid tests, coimmunoprecipitates with Myo5p, and colocalizes with Myo5p. Immunolocalization of the myc-tagged SH3 domain of Myo5p reveals diffuse cytoplasmic staining. Thus, the SH3 domain of Myo5p contributes to but is not sufficient for localization of Myo5p either to patches or to sites of polarized cell growth. Consistent with this, Myo5p patches assemble but do not localize to sites of polarized cell surface growth in a VRP1 deletion mutant. Our studies support a multistep model for Myo5p targeting in yeast. The first step, assembly of Myo5p patches, is dependent upon F-actin, and the second step, polarization of actin patches, requiresVrp1p and the SH3 domain of Myo5p.
出芽酵母含有两种I型肌球蛋白,Myo3p和Myo5p,它们具有冗余功能。两种肌球蛋白的缺失都会导致生长缺陷、肌动蛋白极性丧失和细胞表面极性生长丧失,以及细胞内膜的积累。在myo3Δmyo5Δ细胞中表达myc标签的Myo5p可完全恢复野生型特征。Myo5p定位于点状的皮质结构,在细胞极性生长部位富集。我们发现,latrunculin-A诱导的F-肌动蛋白解聚导致Myo5p斑块的丧失。此外,将酵母细胞在37℃孵育会导致Myo5p斑块和肌动蛋白细胞骨架的短暂去极化。在myo3Δmyo5Δ细胞中表达非运动结构域缺失的突变型Myo5蛋白,并对所得菌株的Myo5p功能进行分析。之前认为与ATP不敏感的肌动蛋白结合有关的尾部同源性2(TH2)结构域的缺失,对Myo5p功能没有可检测到的影响。相反,表达缺失src同源结构域3(SH3)或同时缺失TH2和SH3结构域的突变型Myo5蛋白的myo3Δmyo5Δ细胞表现出缺陷,包括Myo5p斑块去极化、肌动蛋白紊乱以及与肌动蛋白功能障碍相关的表型。这些发现支持SH3结构域在出芽酵母中Myo5p定位和功能中的作用。富含脯氨酸的蛋白verprolin(Vrp1p)在双杂交试验中与Myo3p或Myo5p的SH3结构域结合,与Myo5p共免疫沉淀,并与Myo5p共定位。对Myo5p的myc标签SH3结构域进行免疫定位,显示出弥漫性的细胞质染色。因此,Myo5p的SH3结构域有助于Myo将5p定位到斑块或细胞极性生长部位,但并不足以实现这一定位。与此一致的是,在VRP1缺失突变体中,Myo5p斑块组装但不定位于细胞表面极性生长部位。我们的研究支持酵母中Myo5p靶向的多步骤模型。第一步,Myo5p斑块的组装,依赖于F-肌动蛋白,第二步,肌动蛋白斑块的极化,需要Vrp1p和Myo5p的SH3结构域。