Tiwari-Woodruff S K, Lin M A, Schulteis C T, Papazian D M
Department of Physiology and Molecular Biology Institute, University of California, Los Angeles, School of Medicine, Los Angeles, California 90095-1751, USA.
J Gen Physiol. 2000 Feb;115(2):123-38. doi: 10.1085/jgp.115.2.123.
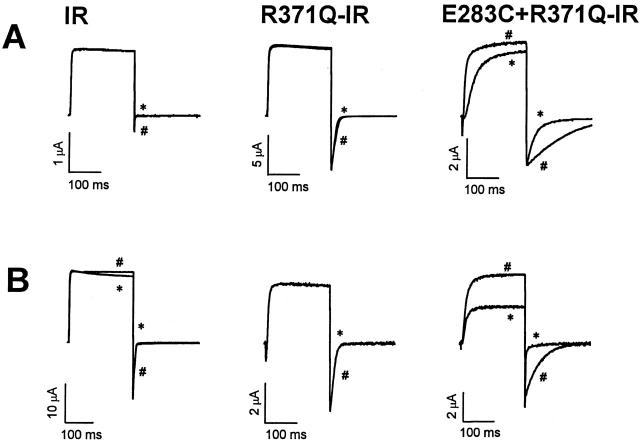
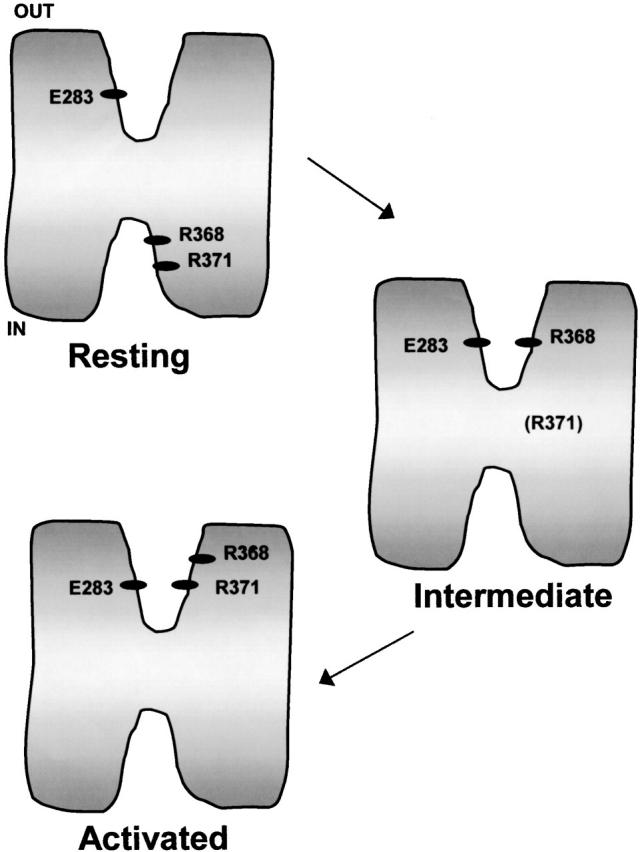
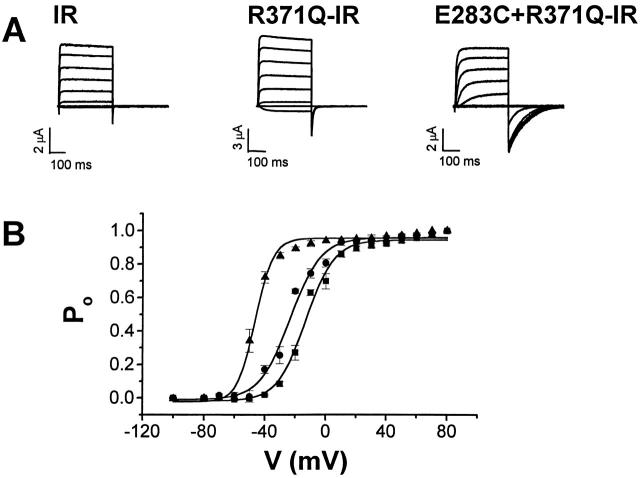
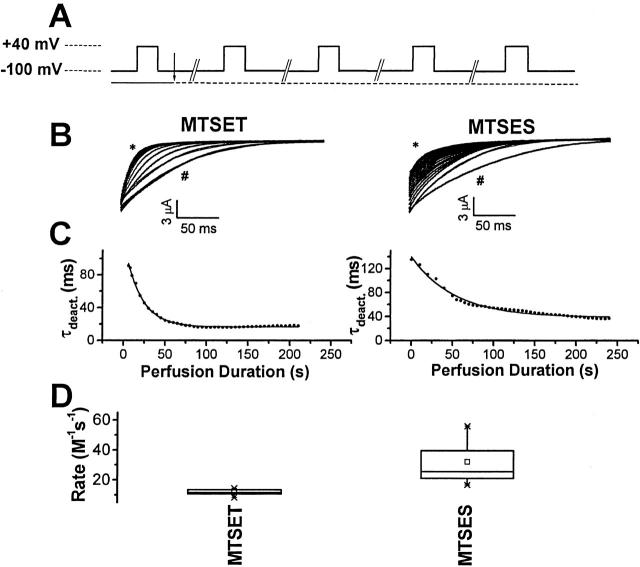
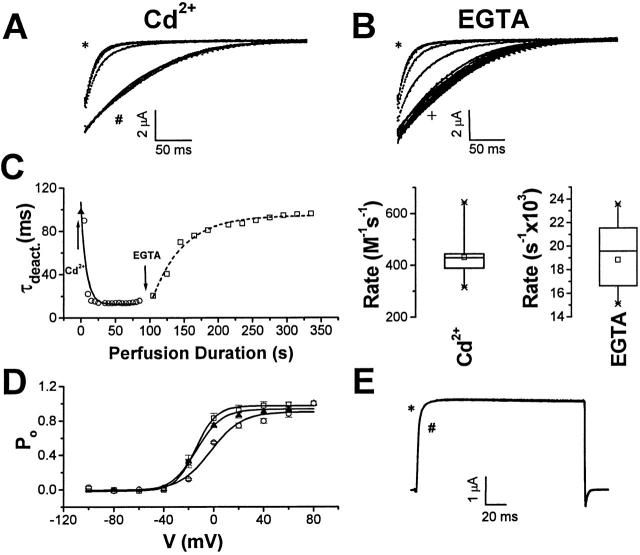
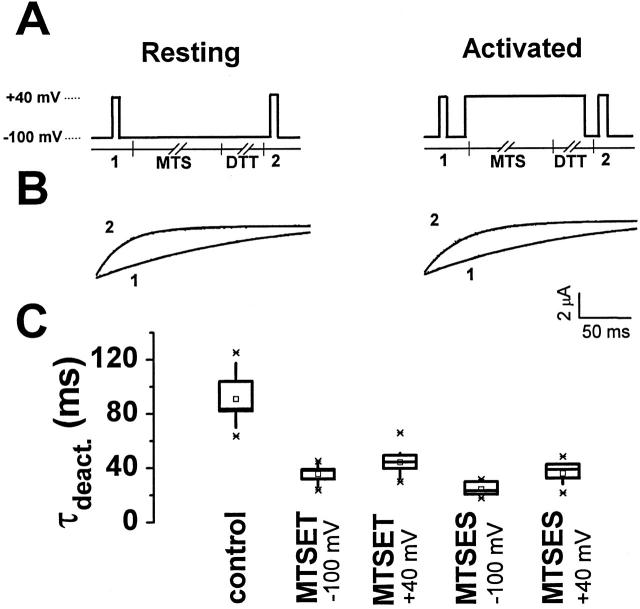
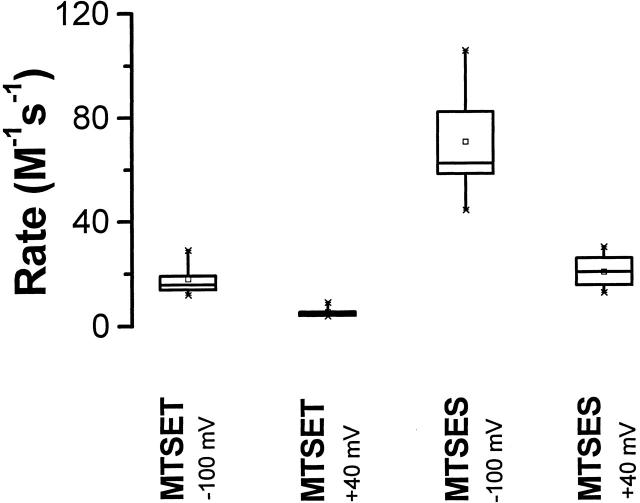
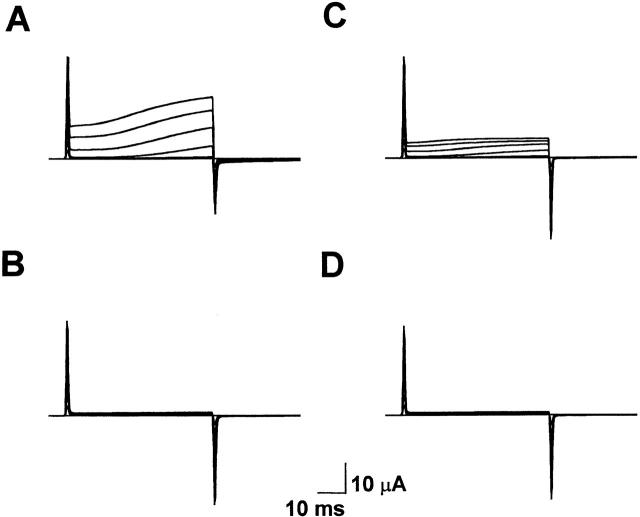
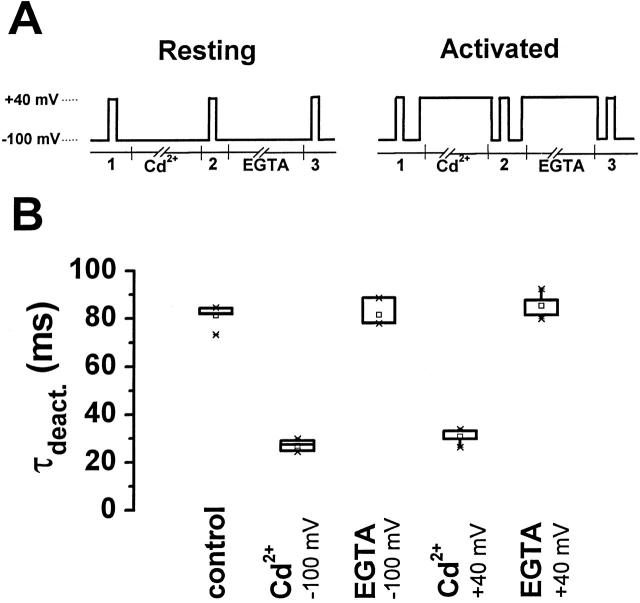
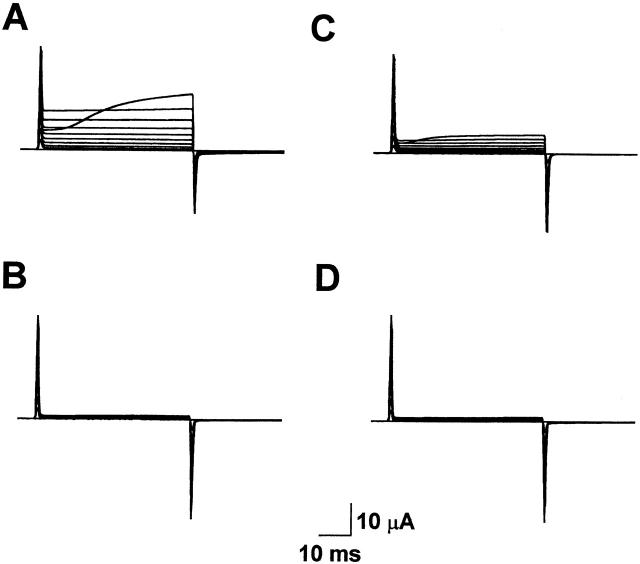
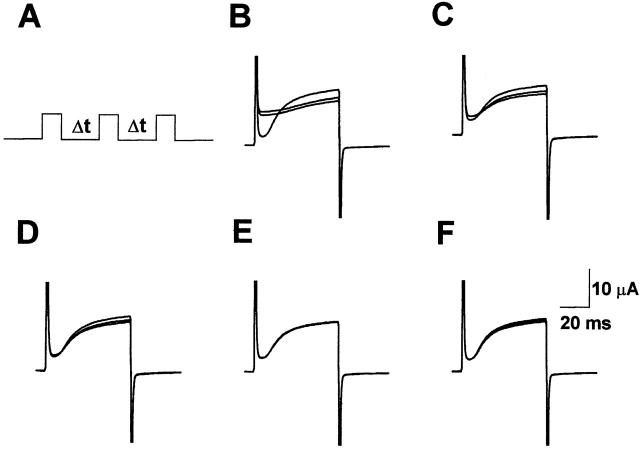
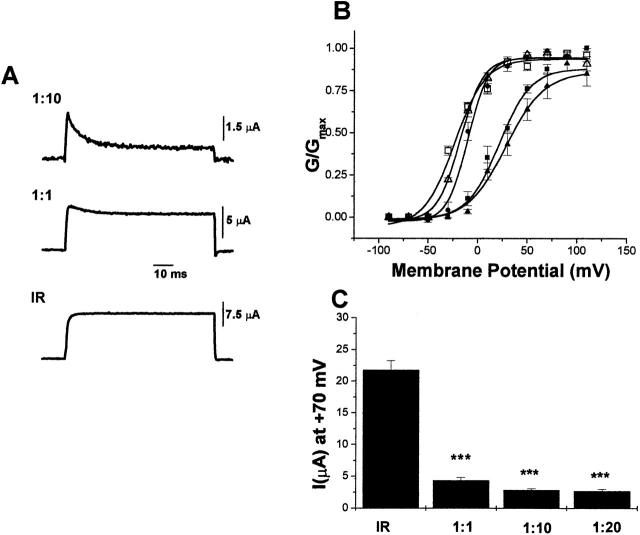
Using a strategy related to intragenic suppression, we previously obtained evidence for structural interactions in the voltage sensor of Shaker K(+) channels between residues E283 in S2 and R368 and R371 in S4 (Tiwari-Woodruff, S.K., C.T. Schulteis, A.F. Mock, and D. M. Papazian. 1997. Biophys. J. 72:1489-1500). Because R368 and R371 are involved in the conformational changes that accompany voltage-dependent activation, we tested the hypothesis that these S4 residues interact with E283 in S2 in a subset of the conformational states that make up the activation pathway in Shaker channels. First, the location of residue 283 at hyperpolarized and depolarized potentials was inferred by substituting a cysteine at that position and determining its reactivity with hydrophilic, sulfhydryl-specific probes. The results indicate that position 283 reacts with extracellularly applied sulfhydryl reagents with similar rates at both hyperpolarized and depolarized potentials. We conclude that E283 is located near the extracellular surface of the protein in both resting and activated conformations. Second, we studied the functional phenotypes of double charge reversal mutations between positions 283 and 368 and between 283 and 371 to gain insight into the conformations in which these positions approach each other most closely. We found that combining charge reversal mutations at positions 283 and 371 stabilized an activated conformation of the channel, and dramatically slowed transitions into and out of this state. In contrast, charge reversal mutations at positions 283 and 368 stabilized a closed conformation, which by virtue of the inferred position of 368 corresponds to a partially activated (intermediate) closed conformation. From these results, we propose a preliminary model for the rearrangement of structural interactions of the voltage sensor during activation of Shaker K(+) channels.
我们之前采用一种与基因内抑制相关的策略,获得了有关Shaker钾通道电压感受器中S2区域的E283残基与S4区域的R368和R371残基之间存在结构相互作用的证据(Tiwari-Woodruff, S.K., C.T. Schulteis, A.F. Mock, and D.M. Papazian. 1997. Biophys. J. 72:1489 - 1500)。由于R368和R371参与了伴随电压依赖性激活的构象变化,我们检验了这样一个假说:在构成Shaker通道激活途径的一部分构象状态中,这些S4残基与S2中的E283相互作用。首先,通过在该位置替换一个半胱氨酸并确定其与亲水性、巯基特异性探针的反应性,推断出残基283在超极化和去极化电位下的位置。结果表明,在超极化和去极化电位下,位置283与细胞外应用的巯基试剂的反应速率相似。我们得出结论,在静息和激活构象中,E283均位于蛋白质的细胞外表面附近。其次,我们研究了283位与368位之间以及283位与371位之间双电荷反转突变的功能表型,以深入了解这些位置彼此最接近时的构象。我们发现,将283位和371位的电荷反转突变组合起来可稳定通道的激活构象,并显著减缓进出该状态的转变。相比之下,283位和368位的电荷反转突变稳定了一种关闭构象,根据推断的368位位置,该构象对应于一种部分激活(中间)的关闭构象。基于这些结果,我们提出了一个关于Shaker钾通道激活过程中电压感受器结构相互作用重排的初步模型。