Gennity J M, Kim H, Inouye M
Department of Biochemistry, University of Medicine and Dentistry of New Jersey, Robert Wood Johnson Medical School, Piscataway 08854-5635.
J Bacteriol. 1992 Apr;174(7):2095-101. doi: 10.1128/jb.174.7.2095-2101.1992.
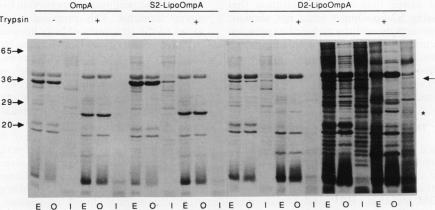
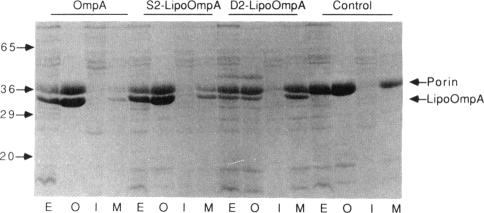
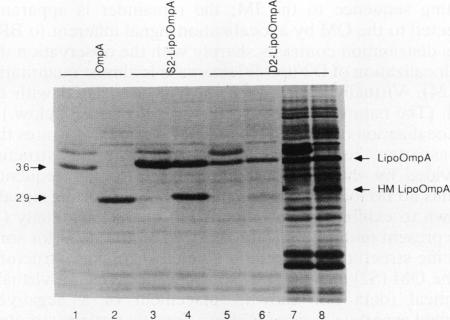
The lipid-modified nine-residue amino-terminal sequence of the mature form of the major outer membrane lipoprotein of Escherichia coli contains information that is responsible for sorting to either the inner or outer membrane. Fusion of this sorting sequence to beta-lactamase is sufficient for localization of the resultant lipo-beta-lactamase to the outer membrane (J. Ghrayeb and M. Inouye, J. Biol. Chem. 259:463-467, 1984). Substitution of the serine adjacent to the amino-terminal lipid-modified cysteine residue of the sorting sequence with the negatively charged residue aspartate causes inner membrane localization (K. Yamaguchi, F. Yu, and M. Inouye, Cell 53:423-432, 1988). Fusion of the aspartate-containing nine-residue inner membrane localization signal to the normally outer membrane lipoprotein bacteriocin release protein does cause partial localization to the inner membrane. However, a single replacement of the glutamine adjacent to the amino-terminal lipid-modified cysteine residue of bacteriocin release protein with aspartate causes no inner membrane localization. Therefore, an aspartate residue itself lacks the information necessary for inner membrane sorting when removed from the structural context provided by the additional eight residues of the sorting sequence. Although the aspartate-containing inner membrane sorting sequence causes an almost quantitative localization to the inner membrane when fused to the otherwise soluble protein beta-lactamase, this sequence cannot prevent significant outer membrane localization when fused to proteins (bacteriocin release protein and OmpA) normally found in the outer membrane. Therefore, structural determinants in addition to the amino-terminal sorting sequence influence the membrane localization of lipoproteins.
大肠杆菌主要外膜脂蛋白成熟形式的脂质修饰九残基氨基末端序列包含负责分选至内膜或外膜的信息。该分选序列与β-内酰胺酶融合足以使所得的脂化β-内酰胺酶定位于外膜(J. Ghrayeb和M. Inouye,《生物化学杂志》259:463 - 467,1984年)。将分选序列中氨基末端脂质修饰的半胱氨酸残基相邻的丝氨酸替换为带负电荷的天冬氨酸残基会导致内膜定位(K. Yamaguchi、F. Yu和M. Inouye,《细胞》53:423 - 432,1988年)。将含天冬氨酸的九残基内膜定位信号与通常定位于外膜的脂蛋白细菌素释放蛋白融合确实会导致部分定位于内膜。然而,将细菌素释放蛋白氨基末端脂质修饰的半胱氨酸残基相邻的谷氨酰胺单一替换为天冬氨酸不会导致内膜定位。因此,当从天冬氨酸分选序列的另外八个残基提供的结构背景中去除时,天冬氨酸残基本身缺乏内膜分选所需的信息。尽管含天冬氨酸的内膜分选序列与原本可溶的蛋白β-内酰胺酶融合时会导致几乎定量地定位于内膜,但该序列与通常在外膜中发现的蛋白(细菌素释放蛋白和OmpA)融合时不能阻止显著的外膜定位。因此,除了氨基末端分选序列外,结构决定因素也影响脂蛋白的膜定位。