Russell Vivienne A, Oades Robert D, Tannock Rosemary, Killeen Peter R, Auerbach Judith G, Johansen Espen B, Sagvolden Terje
Department of Human Biology, Faculty of Health Sciences, University of Cape Town, Anzio Road, Observatory 7925, South Africa.
Behav Brain Funct. 2006 Aug 23;2:30. doi: 10.1186/1744-9081-2-30.
Current concepts of Attention-Deficit/Hyperactivity Disorder (ADHD) emphasize the role of higher-order cognitive functions and reinforcement processes attributed to structural and biochemical anomalies in cortical and limbic neural networks innervated by the monoamines, dopamine, noradrenaline and serotonin. However, these explanations do not account for the ubiquitous findings in ADHD of intra-individual performance variability, particularly on tasks that require continual responses to rapid, externally-paced stimuli. Nor do they consider attention as a temporal process dependent upon a continuous energy supply for efficient and consistent function. A consideration of this feature of intra-individual response variability, which is not unique to ADHD but is also found in other disorders, leads to a new perspective on the causes and potential remedies of specific aspects of ADHD.
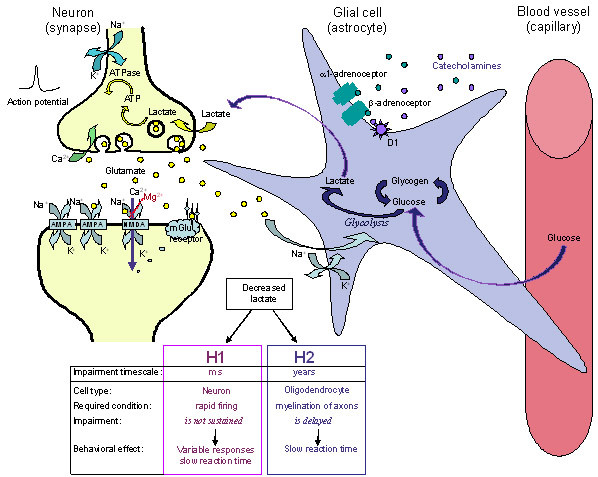
We propose that in ADHD, astrocyte function is insufficient, particularly in terms of its formation and supply of lactate. This insufficiency has implications both for performance and development: H1) In rapidly firing neurons there is deficient ATP production, slow restoration of ionic gradients across neuronal membranes and delayed neuronal firing; H2) In oligodendrocytes insufficient lactate supply impairs fatty acid synthesis and myelination of axons during development. These effects occur over vastly different time scales: those due to deficient ATP (H1) occur over milliseconds, whereas those due to deficient myelination (H2) occur over months and years. Collectively the neural outcomes of impaired astrocytic release of lactate manifest behaviourally as inefficient and inconsistent performance (variable response times across the lifespan, especially during activities that require sustained speeded responses and complex information processing).
Multi-level and multi-method approaches are required. These include: 1) Use of dynamic strategies to evaluate cognitive performance under conditions that vary in duration, complexity, speed, and reinforcement; 2) Use of sensitive neuroimaging techniques such as diffusion tensor imaging, magnetic resonance spectroscopy, electroencephalography or magnetoencephalopathy to quantify developmental changes in myelination in ADHD as a potential basis for the delayed maturation of brain function and coordination, and 3) Investigation of the prevalence of genetic markers for factors that regulate energy metabolism (lactate, glutamate, glucose transporters, glycogen synthase, glycogen phosphorylase, glycolytic enzymes), release of glutamate from synaptic terminals and glutamate-stimulated lactate production (SNAP25, glutamate receptors, adenosine receptors, neurexins, intracellular Ca2+), as well as astrocyte function (alpha1, alpha2 and beta-adrenoceptors, dopamine D1 receptors) and myelin synthesis (lactate transporter, Lingo-1, Quaking homolog, leukemia inhibitory factor, and Transferrin).
The hypothesis extends existing theories of ADHD by proposing a physiological basis for specific aspects of the ADHD phenotype - namely frequent, transient and impairing fluctuations in functioning, particularly during performance of speeded, effortful tasks. The immediate effects of deficient ATP production and slow restoration of ionic gradients across membranes of rapidly firing neurons have implications for daily functioning: For individuals with ADHD, performance efficacy would be enhanced if repetitive and lengthy effortful tasks were segmented to reduce concurrent demands for speed and accuracy of response (introduction of breaks into lengthy/effortful activities such as examinations, motorway driving, assembly-line production). Also, variations in task or modality and the use of self- rather than system-paced schedules would be helpful. This would enable energetic demands to be distributed to alternate neural resources, and energy reserves to be re-established. Longer-term effects may manifest as reduction in regional brain volumes since brain areas with the highest energy demand will be most affected by a restricted energy supply and may be reduced in size. Novel forms of therapeutic agent and delivery system could be based on factors that regulate energy production and myelin synthesis. Since the phenomena and our proposed basis for it are not unique to ADHD but also manifests in other disorders, the implications of our hypotheses may be relevant to understanding and remediating these other conditions as well.
当前对注意力缺陷多动障碍(ADHD)的认识强调高阶认知功能以及归因于单胺类、多巴胺、去甲肾上腺素和血清素支配的皮质和边缘神经网络中结构和生化异常的强化过程的作用。然而,这些解释无法说明ADHD中普遍存在的个体内表现变异性,特别是在需要对快速的外部节奏刺激持续做出反应的任务中。它们也没有将注意力视为一个依赖持续能量供应以实现高效和一致功能的时间过程。考虑到这种个体内反应变异性的特征,它并非ADHD所特有,在其他疾病中也存在,这为ADHD特定方面的病因及潜在治疗方法带来了新的视角。
我们提出,在ADHD中,星形胶质细胞功能不足,特别是在乳酸的生成和供应方面。这种不足对表现和发育都有影响:H1)在快速放电的神经元中,ATP生成不足,跨神经元膜的离子梯度恢复缓慢,神经元放电延迟;H2)在少突胶质细胞中,乳酸供应不足会损害发育过程中轴突的脂肪酸合成和髓鞘形成。这些影响发生在截然不同的时间尺度上:由于ATP不足(H1)导致的影响发生在毫秒级,而由于髓鞘形成不足(H2)导致的影响发生在数月和数年。星形胶质细胞释放乳酸受损的综合神经结果在行为上表现为低效和不一致的表现(整个生命周期中反应时间可变,尤其是在需要持续快速反应和复杂信息处理的活动中)。
需要采用多层次和多方法的途径。这些包括:1)使用动态策略在持续时间、复杂性、速度和强化程度不同的条件下评估认知表现;2)使用敏感的神经成像技术,如扩散张量成像、磁共振波谱、脑电图或脑磁图,以量化ADHD中髓鞘形成的发育变化,作为脑功能和协调性延迟成熟的潜在基础;3)研究调节能量代谢(乳酸、谷氨酸、葡萄糖转运体、糖原合酶、糖原磷酸化酶、糖酵解酶)、突触末端谷氨酸释放和谷氨酸刺激的乳酸生成(SNAP25、谷氨酸受体、腺苷受体、神经连接蛋白、细胞内Ca2+)以及星形胶质细胞功能(α1、α2和β肾上腺素能受体、多巴胺D1受体)和髓鞘合成(乳酸转运体、Lingo-1、震颤同源物、白血病抑制因子和转铁蛋白)的因素的遗传标记的患病率。
该假说通过为ADHD表型的特定方面提出生理基础,扩展了现有的ADHD理论——即功能频繁、短暂且有害的波动,特别是在快速、费力任务的执行过程中。快速放电神经元中ATP生成不足以及跨膜离子梯度恢复缓慢的直接影响对日常功能有影响:对于ADHD患者,如果将重复性和冗长的费力任务进行分段,以减少对反应速度和准确性的同时需求(在冗长/费力的活动如考试、高速公路驾驶、流水线生产中引入休息),表现效能将会提高。此外,任务或方式的变化以及使用自我节奏而非系统节奏的时间表会有所帮助。这将使能量需求能够分配到交替的神经资源上,并重新建立能量储备。长期影响可能表现为局部脑容量减少,因为能量需求最高的脑区将最受能量供应受限的影响,可能会缩小。新型治疗药物和给药系统可以基于调节能量生成和髓鞘合成的因素。由于这些现象以及我们提出的其基础并非ADHD所特有,在其他疾病中也有体现,我们假说的意义可能也与理解和治疗这些其他疾病相关。