Liu Andrew C, Welsh David K, Ko Caroline H, Tran Hien G, Zhang Eric E, Priest Aaron A, Buhr Ethan D, Singer Oded, Meeker Kirsten, Verma Inder M, Doyle Francis J, Takahashi Joseph S, Kay Steve A
Department of Biochemistry, The Scripps Research Institute, La Jolla, CA 92037, USA.
Cell. 2007 May 4;129(3):605-16. doi: 10.1016/j.cell.2007.02.047.
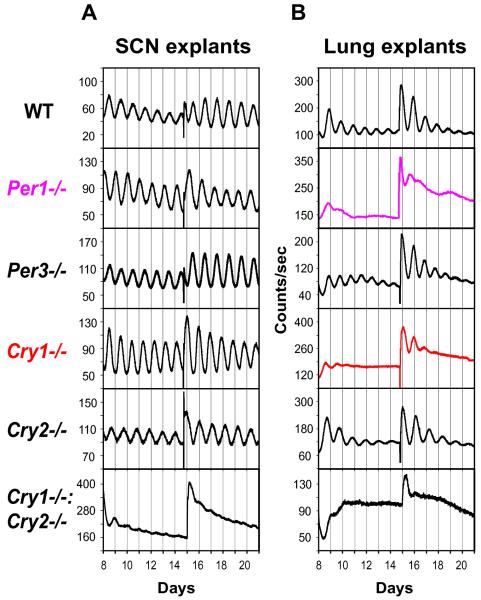
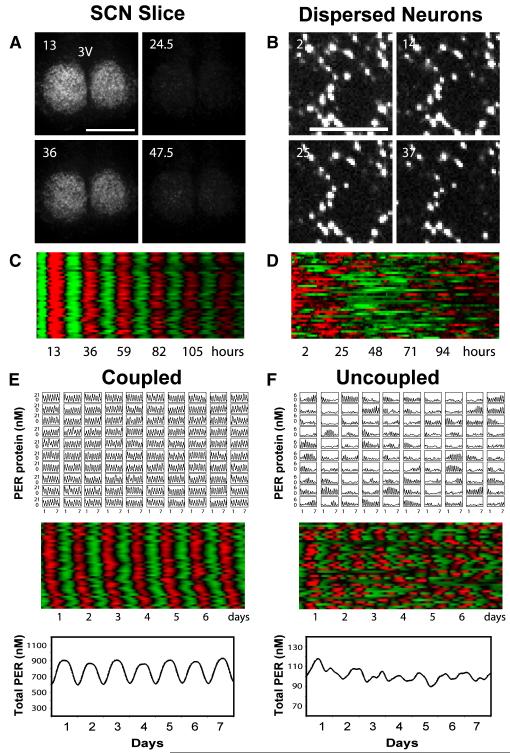
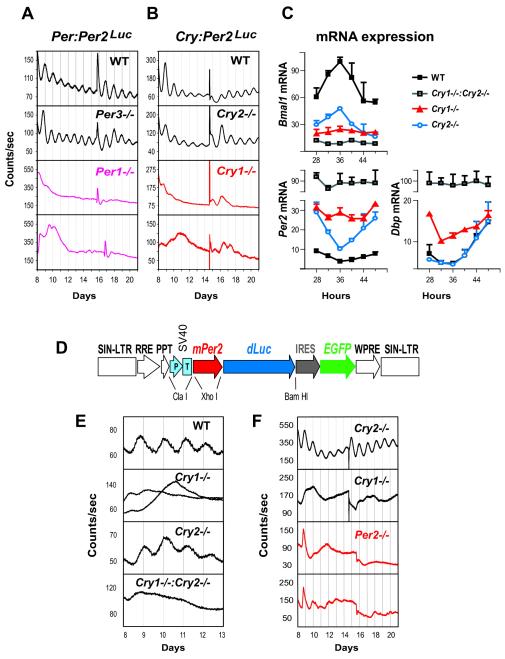
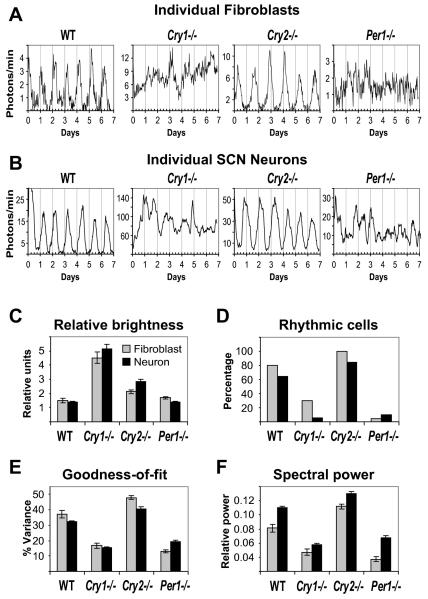
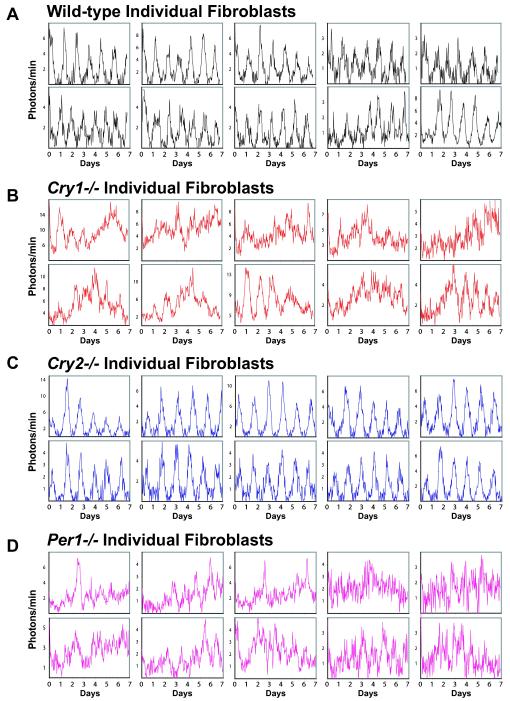
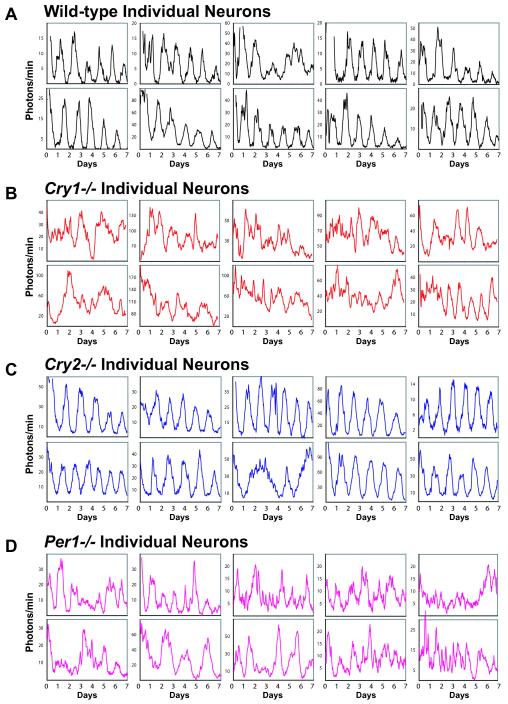
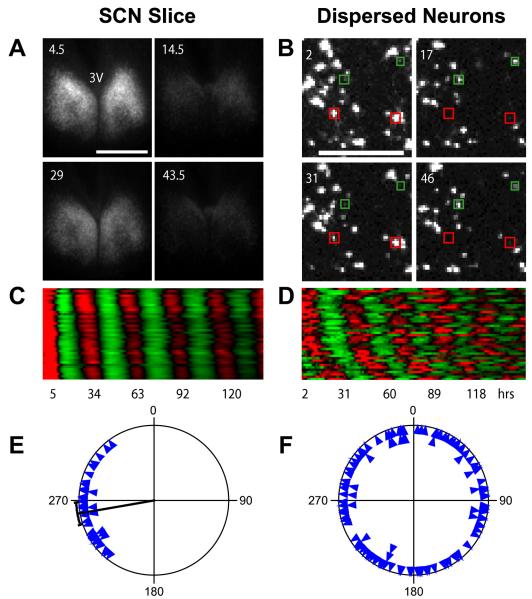
Molecular mechanisms of the mammalian circadian clock have been studied primarily by genetic perturbation and behavioral analysis. Here, we used bioluminescence imaging to monitor Per2 gene expression in tissues and cells from clock mutant mice. We discovered that Per1 and Cry1 are required for sustained rhythms in peripheral tissues and cells, and in neurons dissociated from the suprachiasmatic nuclei (SCN). Per2 is also required for sustained rhythms, whereas Cry2 and Per3 deficiencies cause only period length defects. However, oscillator network interactions in the SCN can compensate for Per1 or Cry1 deficiency, preserving sustained rhythmicity in mutant SCN slices and behavior. Thus, behavior does not necessarily reflect cell-autonomous clock phenotypes. Our studies reveal previously unappreciated requirements for Per1, Per2, and Cry1 in sustaining cellular circadian rhythmicity and demonstrate that SCN intercellular coupling is essential not only to synchronize component cellular oscillators but also for robustness against genetic perturbations.
哺乳动物生物钟的分子机制主要通过基因扰动和行为分析进行研究。在此,我们利用生物发光成像技术监测生物钟突变小鼠组织和细胞中的Per2基因表达。我们发现,Per1和Cry1是外周组织、细胞以及从视交叉上核(SCN)分离出的神经元中维持节律所必需的。Per2对于维持节律也是必需的,而Cry2和Per3缺陷仅导致周期长度缺陷。然而,SCN中的振荡器网络相互作用可以补偿Per1或Cry1缺陷,在突变的SCN切片和行为中保持持续的节律性。因此,行为不一定反映细胞自主的生物钟表型。我们的研究揭示了Per1、Per2和Cry1在维持细胞昼夜节律性方面以前未被认识到的需求,并证明SCN细胞间耦合不仅对于同步组成细胞振荡器至关重要,而且对于抵抗基因扰动的稳健性也至关重要。