Department of Psychological and Brain Sciences, Center for Cognitive Neuroscience, Dartmouth College, Hanover, New Hampshire, USA.
Hippocampus. 2011 Jul;21(7):767-82. doi: 10.1002/hipo.20874. Epub 2010 Nov 3.
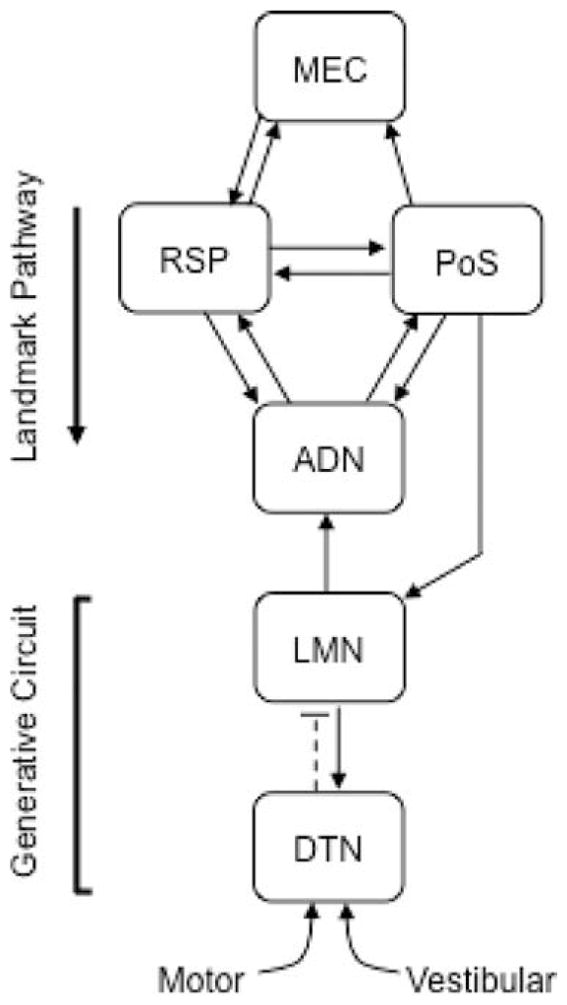
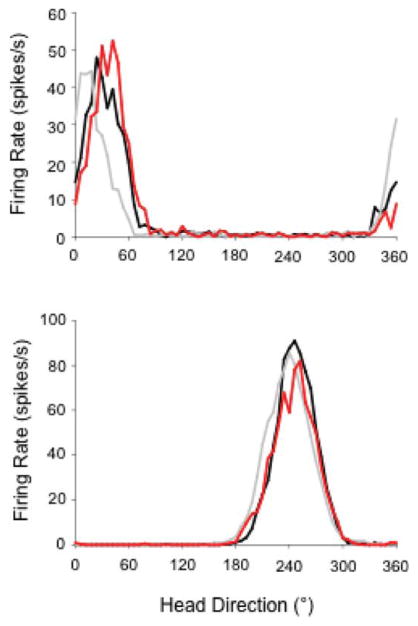
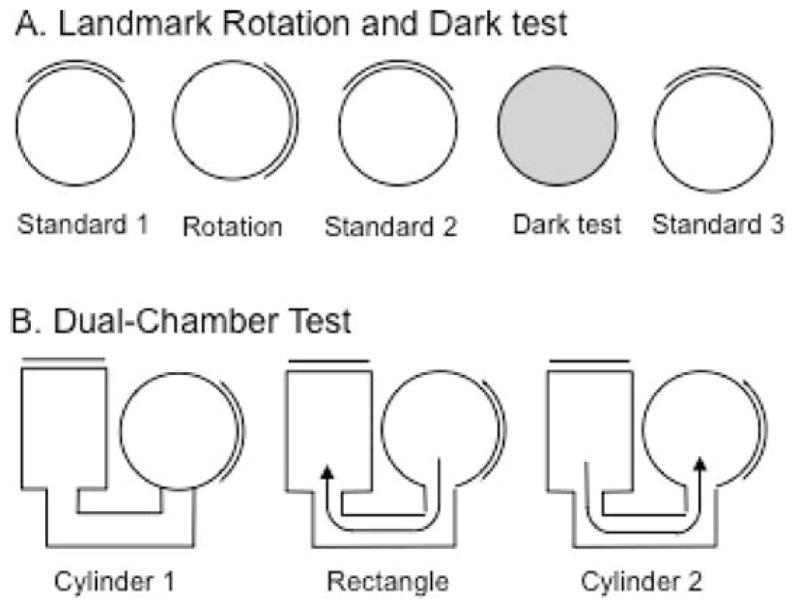
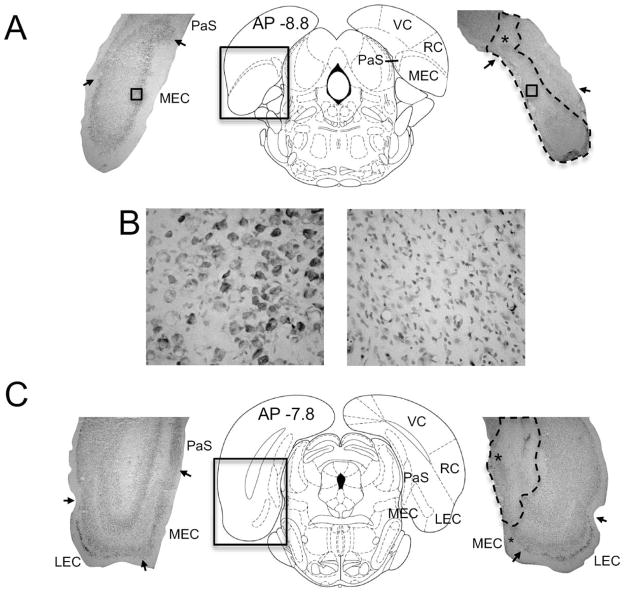
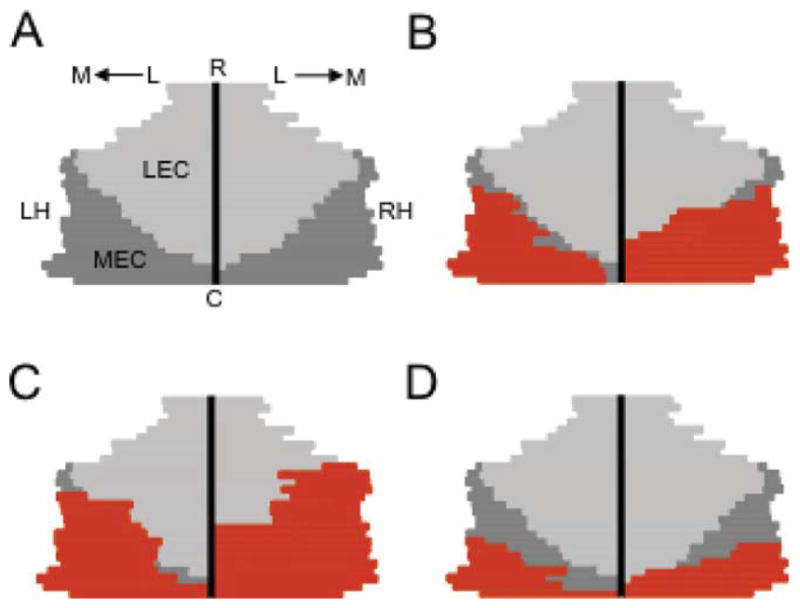
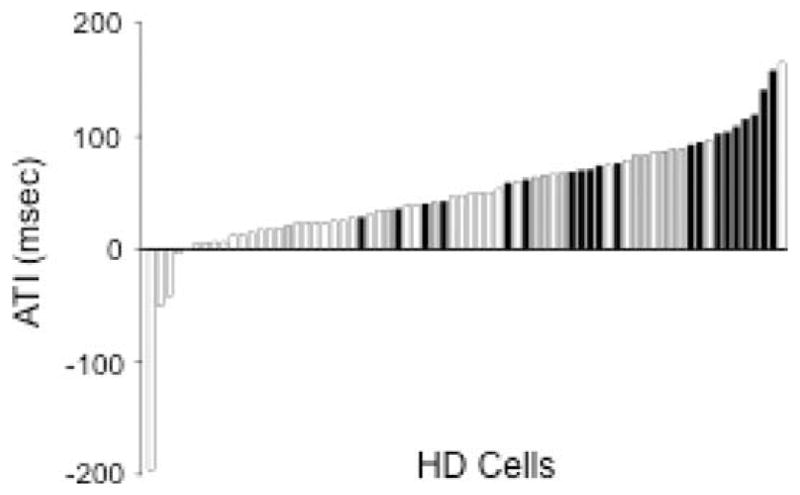
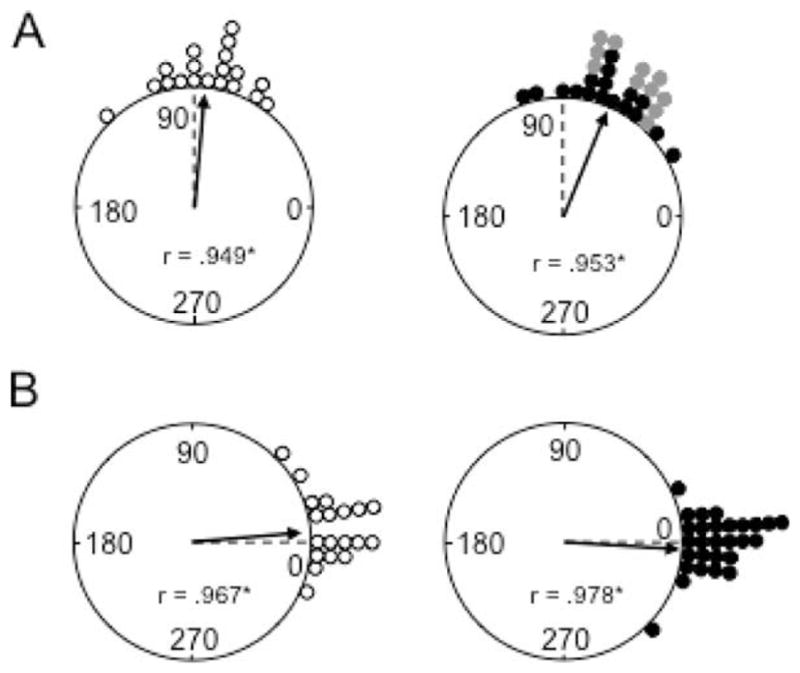
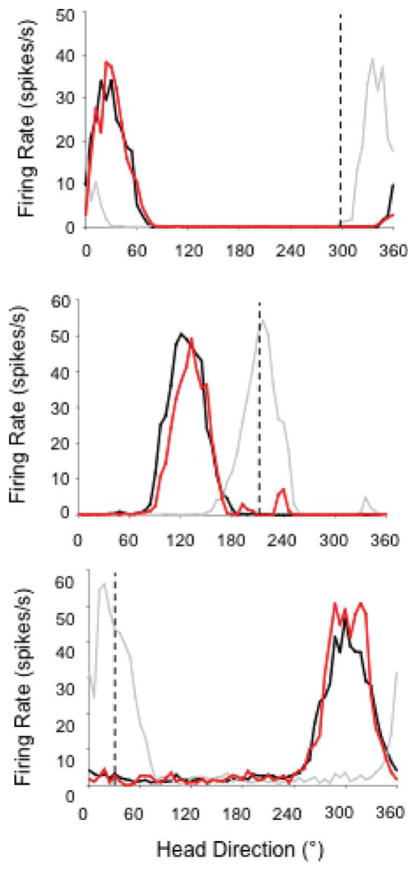
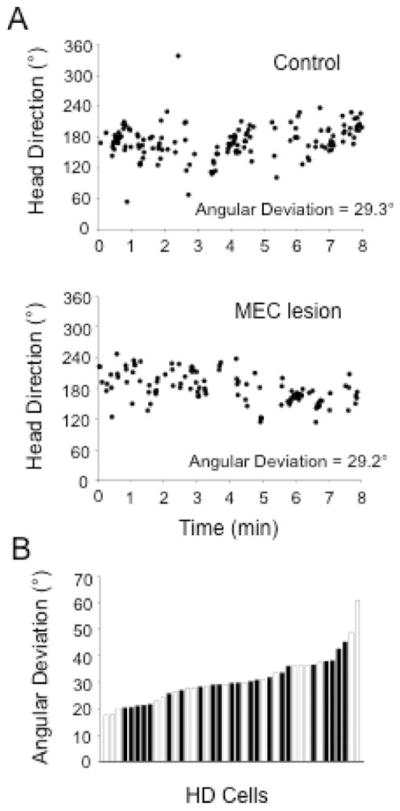
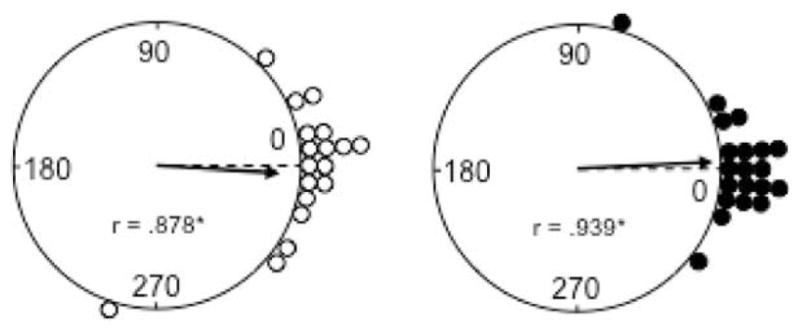
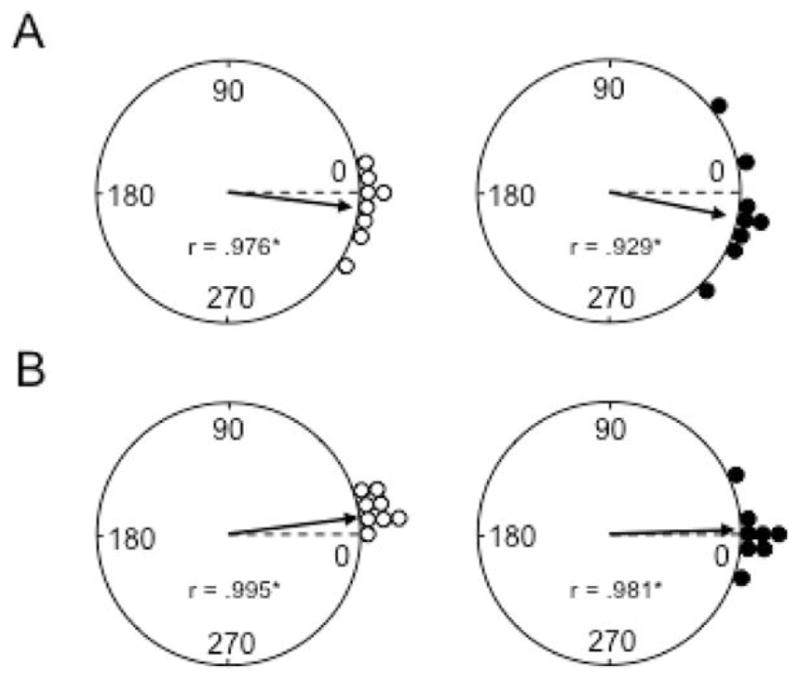
The medial entorhinal cortex (MEC) occupies a central position within neural circuits devoted to the representation of spatial location and orientation. The MEC contains cells that fire as a function of the animal's head direction (HD), as well as grid cells that fire in multiple locations in an environment, forming a repeating hexagonal pattern. The MEC receives inputs from widespread areas of the cortical mantle including the ventral visual stream, which processes object recognition information, as well as information about visual landmarks. The role of the MEC in processing the HD signal or landmark information is unclear. We addressed this issue by neurotoxically damaging the MEC and recording HD cells within the anterodorsal thalamus (ADN). Direction-specific activity was present in the ADN of all animals with MEC lesions. Moreover, the discharge characteristics of ADN HD cells were only mildly affected by MEC lesions, with HD cells exhibiting greater anticipation of future HDs. Tests of landmark control revealed that HD cells in lesioned rats were capable of accurately updating their preferred firing directions in relation to a salient visual cue. Furthermore, cells from lesioned animals maintained stable preferred firing directions when locomoting in darkness and demonstrated stable HD cell tuning when locomoting into a novel enclosure, suggesting that MEC lesions did not disrupt the integration of idiothetic cues, or angular path integration, by HD cells. Collectively, these findings suggest that the MEC plays a limited role in the formation and spatial updating of the HD cell signal.
内侧缰状回(MEC)在专门用于表示空间位置和方向的神经回路中占据中心位置。MEC 包含随动物头部方向(HD)发射的细胞,以及在环境中的多个位置发射的网格细胞,形成重复的六边形模式。MEC 接收来自皮质帽广泛区域的输入,包括处理物体识别信息的腹侧视觉流,以及关于视觉地标信息。MEC 在处理 HD 信号或地标信息方面的作用尚不清楚。我们通过神经毒素损伤 MEC 并在背侧前丘脑(ADN)中记录 HD 细胞来解决这个问题。所有 MEC 损伤动物的 ADN 中都存在方向特异性活动。此外,ADN HD 细胞的放电特性仅受到 MEC 损伤的轻微影响,HD 细胞表现出对未来 HD 的更大预期。地标控制测试表明,损伤大鼠的 HD 细胞能够准确地根据显著的视觉线索更新其首选发射方向。此外,来自损伤动物的细胞在黑暗中运动时保持稳定的首选发射方向,并在进入新围场时表现出稳定的 HD 细胞调谐,这表明 MEC 损伤不会破坏 HD 细胞对自身感觉线索或角路径整合的整合。总的来说,这些发现表明 MEC 在 HD 细胞信号的形成和空间更新中仅发挥有限作用。