Max Planck Institute for Developmental Biology, Spemannstrasse 35, 72076 Tübingen, Germany.
J Biol Chem. 2011 Feb 25;286(8):6280-90. doi: 10.1074/jbc.M110.165274. Epub 2010 Dec 13.
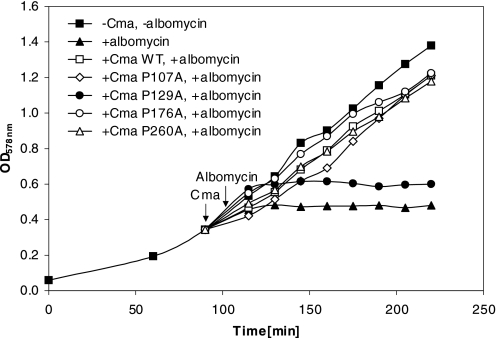
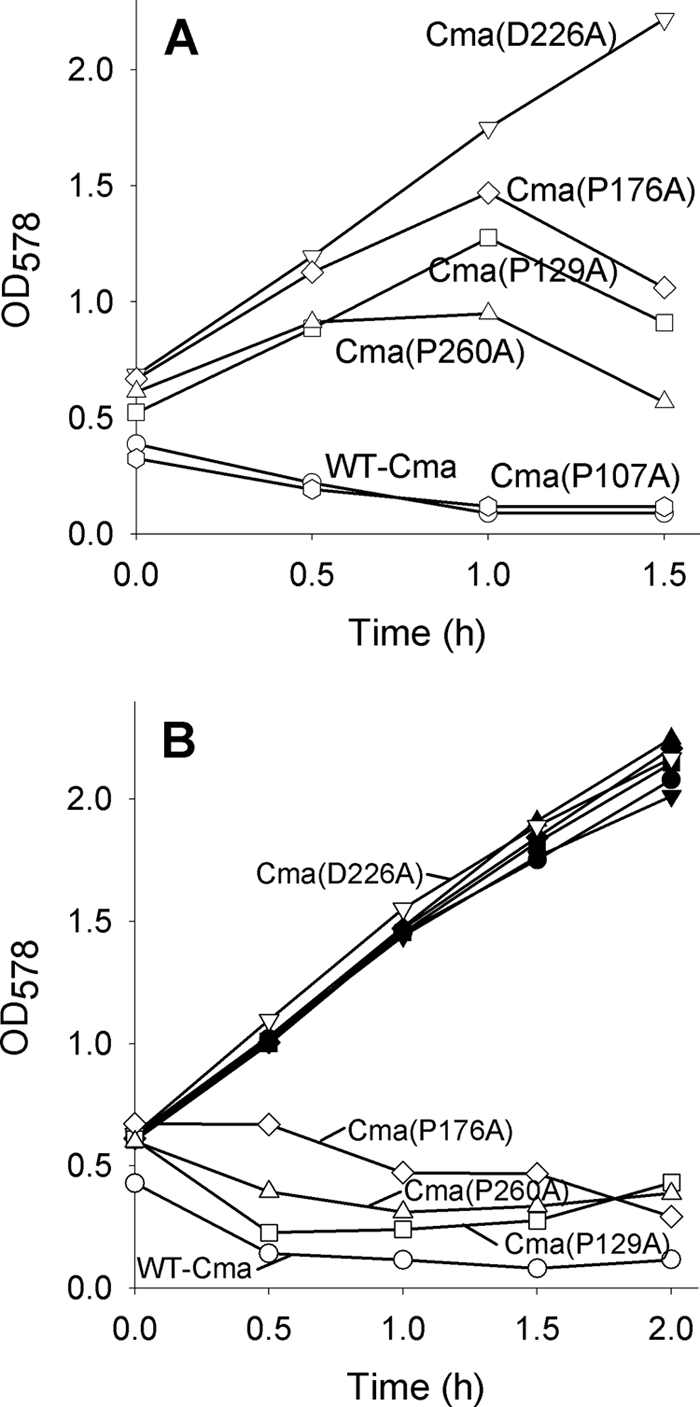
Colicin M (Cma) is specifically imported into the periplasm of Escherichia coli and kills the cells. Killing depends on the periplasmic peptidyl prolyl cis-trans isomerase/chaperone FkpA. To identify the Cma prolyl bonds targeted by FkpA, we replaced the 15 proline residues individually with alanine. Seven mutant proteins were fully active; Cma(P129A), Cma(P176A), and Cma(P260A) displayed 1%, and Cma(P107A) displayed 10% of the wild-type activity. Cma(P107A), Cma(P129A), and Cma(P260A), but not Cma(P176A), killed cells after entering the periplasm via osmotic shock, indicating that the former mutants were translocation-deficient; Cma(P129A) did not bind to the FhuA outer membrane receptor. The crystal structures of Cma and Cma(P176A) were identical, excluding inactivation of the activity domain located far from Pro-176. In a new peptidyl prolyl cis-trans isomerase assay, FkpA isomerized the Cma prolyl bond in peptide Phe-Pro-176 at a high rate, but Lys-Pro-107 and Leu-Pro-260 isomerized at only <10% of that rate. The four mutant proteins secreted into the periplasm via a fused signal sequence were toxic but much less than wild-type Cma. Wild-type and mutant Cma proteins secreted or translocated across the outer membrane by energy-coupled import or unspecific osmotic shock were only active in the presence of FkpA. We propose that Cma unfolds during transfer across the outer or cytoplasmic membrane and refolds to the active form in the periplasm assisted by FkpA. Weak refolding of Cma(P176A) would explain its low activity in all assays. Of the four proline residues identified as being important for Cma activity, Phe-Pro-176 is most likely targeted by FkpA.
大肠杆菌素 M(Cma)特异性地被导入大肠杆菌的周质并杀死细胞。这种杀伤依赖于周质的肽基脯氨酰顺反异构酶/伴侣 FkpA。为了鉴定 FkpA 靶向的 Cma 脯氨酸键,我们将 15 个脯氨酸残基逐个突变为丙氨酸。七种突变蛋白均具有完全活性;Cma(P129A)、Cma(P176A)和 Cma(P260A)的活性分别为野生型的 1%和 10%,而 Cma(P107A)的活性为野生型的 10%。Cma(P107A)、Cma(P129A)和 Cma(P260A),但不是 Cma(P176A),在通过渗透压休克进入周质后能够杀死细胞,表明前三种突变体易位缺陷;Cma(P129A)不能与 FhuA 外膜受体结合。Cma 和 Cma(P176A)的晶体结构相同,排除了位于 Pro-176 远端的活性结构域失活。在新的肽基脯氨酰顺反异构酶测定中,FkpA 以高速度异构化肽 Phe-Pro-176 中的 Cma 脯氨酸键,但 Lys-Pro-107 和 Leu-Pro-260 仅以该速率的<10%异构化。通过融合信号序列分泌到周质中的四种突变蛋白是有毒的,但远低于野生型 Cma。通过能量偶联导入或非特异性渗透压休克穿过外膜分泌或易位的野生型和突变型 Cma 蛋白仅在存在 FkpA 的情况下才具有活性。我们提出 Cma 在穿过外膜或细胞质膜转移过程中展开,并在 FkpA 的辅助下在周质中重新折叠为活性形式。Cma(P176A)的弱重折叠可以解释其在所有测定中的低活性。在鉴定出对 Cma 活性重要的四个脯氨酸残基中,Phe-Pro-176 最有可能是 FkpA 的靶标。