Kanikarla-Marie Preeti, Ronald Sharon, De Benedetti Arrigo
Department of Biochemistry and Molecular Biology and the Feist-Weiller Cancer Center, Louisiana State University Health Sciences Center, Shreveport, LA 71130, USA.
BMC Res Notes. 2011 Jan 21;4:13. doi: 10.1186/1756-0500-4-13.
The S. cerevisiae mating type switch model of double-strand break (DSB) repair, utilizing the HO endonuclease, is one of the best studied systems for both Homologous Recombination Repair (HRR) and direct ends-joining repair (Non-Homologous Ends Joining - NHEJ). We have recently transposed that system to a mammalian cell culture model taking advantage of an adenovirus expressing HO and an integrated genomic target. This made it possible to compare directly the mechanism of repair between yeast and mammalian cells for the same type of induced DSB. Studies of DSB repair have emphasized commonality of features, proteins and machineries between organisms, and differences when conservation is not found. Two proteins that stand out that differ between yeast and mammalian cells are DNA-PK, a protein kinase that is activated by the presence of DSBs, and Artemis, a nuclease whose activity is modulated by DNA-PK and ATM. In this report we describe how these two proteins may be involved in a specific pattern of ends-processing at the DSB, particularly in the context of heterochromatin.
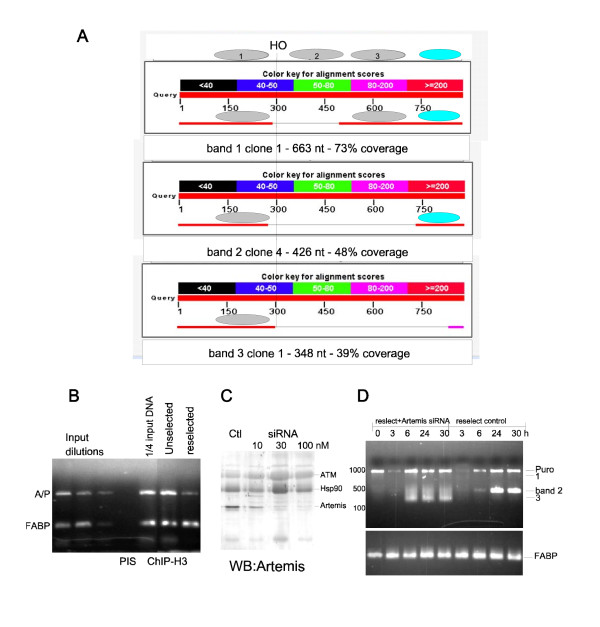
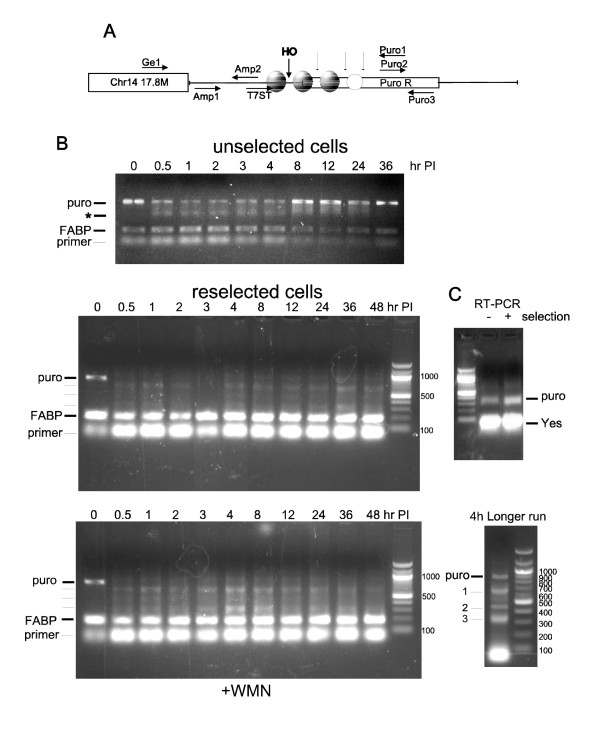
We previously published that the repair of the HO-induced DSB was generally accurate and occurred by simple rejoining of the cohesive 3'-overhangs generated by HO. During continuous passage of those cells in the absence of puromycin selection, the locus appears to have become more heterochromatic and silenced by displaying several features. 1) The site had become less accessible to cleavage by the HO endonuclease; 2) the expression of the puro mRNA, which confers resistance to puromycin, had become reduced; 3) occupancy of nucleosomes at the site (ChIP for histone H3) was increased, an indicator for more condensed chromatin. After reselection of these cells by addition of puromycin, many of these features were reversed. However, even the reselected cells were not identical in the pattern of cleavage and repair as the cells when originally created. Specifically, the pattern of repair revealed discrete deletions at the DSB that indicated unit losses of nucleosomes (or other protein complexes) before religation, represented by a ladder of PCR products reminiscent of an internucleosomal cleavage that is typically observed during apoptosis. This pattern of cleavage suggested to us that perhaps, Artemis, a protein that is believed to generate the internucleosomal fragments during apoptosis and in DSB repair, was involved in that specific pattern of ends-processing. Preliminary evidence indicates that this may be the case, since knock-down of Artemis with siRNA eliminated the laddering pattern and revealed instead an extensive exonucleolytic processing of the ends before religation.
e have generated a system in mammalian cells where the absence of positive selection resulted in chromatin remodeling at the target locus that recapitulates many of the features of the mating-type switching system in yeast. Specifically, just as for yeast HML and HMR, the locus had become transcriptionally repressed; accessibility to cleavage by the HO endonuclease was reduced; and processing of the ends was drastically changed. The switch was from high-fidelity religation of the cohesive ends, to a pattern of release of internucleosomal fragments, perhaps in search of micro-homology stretches for ligation. This is consistent with reports that the involvement of ATM, DNA-PK and Artemis in DSB repair is largely focused to heterochromatic regions, and not required for the majority of IR-induced DSB repair foci in euchromatin.
利用 HO 核酸内切酶的酿酒酵母交配型转换双链断裂(DSB)修复模型,是同源重组修复(HRR)和直接末端连接修复(非同源末端连接 - NHEJ)研究得最为深入的系统之一。我们最近利用表达 HO 的腺病毒和整合的基因组靶点,将该系统转换到了哺乳动物细胞培养模型中。这使得直接比较酵母和哺乳动物细胞中相同类型诱导 DSB 的修复机制成为可能。DSB 修复研究强调了不同生物体之间特征、蛋白质和机制的共性,以及未发现保守性时的差异。酵母和哺乳动物细胞之间显著不同的两种蛋白质是 DNA-PK(一种由 DSB 激活的蛋白激酶)和 Artemis(一种其活性受 DNA-PK 和 ATM 调节的核酸酶)。在本报告中,我们描述了这两种蛋白质可能如何参与 DSB 处特定的末端加工模式,特别是在异染色质的背景下。
我们之前发表过,HO 诱导的 DSB 的修复通常是准确的,通过 HO 产生的粘性 3' 突出端的简单重新连接而发生。在那些细胞在没有嘌呤霉素选择的情况下连续传代过程中,该位点似乎变得更加异染色质化并通过表现出几个特征而沉默。1)该位点对 HO 核酸内切酶的切割变得更难接近;2)赋予对嘌呤霉素抗性的嘌呤霉素 mRNA 的表达降低;3)该位点核小体的占据(组蛋白 H3 的染色质免疫沉淀)增加,这是染色质更浓缩的一个指标。通过添加嘌呤霉素重新选择这些细胞后,许多这些特征都被逆转。然而,即使是重新选择的细胞在切割和修复模式上也与最初创建时的细胞不同。具体而言,修复模式显示 DSB 处有离散的缺失,这表明在重新连接之前核小体(或其他蛋白质复合物)的单位丢失,由一系列 PCR 产物表示,让人联想到通常在细胞凋亡期间观察到的核小体间切割。这种切割模式向我们表明,也许 Artemis(一种被认为在细胞凋亡和 DSB 修复过程中产生核小体间片段的蛋白质)参与了那种特定的末端加工模式。初步证据表明可能是这种情况,因为用 siRNA 敲低 Artemis 消除了梯状模式,取而代之的是在重新连接之前末端的广泛外切核酸酶加工。
我们在哺乳动物细胞中建立了一个系统,其中缺乏阳性选择导致靶位点的染色质重塑,重现了酵母交配型转换系统的许多特征。具体而言,就像酵母的 HML 和 HMR 一样,该位点转录受到抑制;HO 核酸内切酶的切割可及性降低;末端加工发生了巨大变化。转变是从粘性末端的高保真重新连接,到核小体间片段释放的模式,也许是为了寻找微同源序列进行连接。这与 ATM、DNA-PK 和 Artemis 参与 DSB 修复主要集中在异染色质区域的报道一致,并且对于常染色质中大多数 IR 诱导的 DSB 修复位点不是必需的。