Centro de Investigaciones en Química Biológica de Córdoba, CIQUIBIC (UNC-CONICET), Departamento de Química Biológica, Facultad de Ciencias Químicas, Universidad Nacional de Córdoba, Córdoba, Argentina.
PLoS One. 2011 Feb 24;6(2):e16969. doi: 10.1371/journal.pone.0016969.
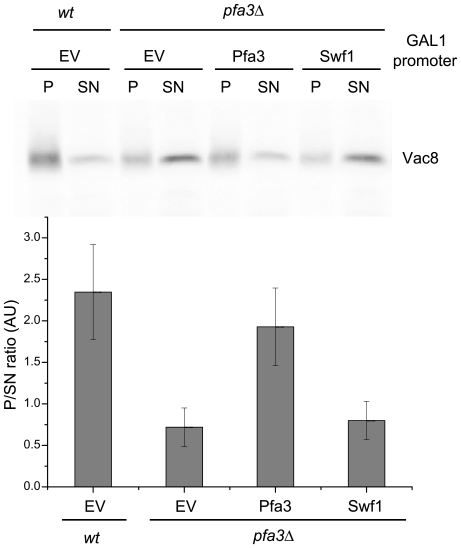
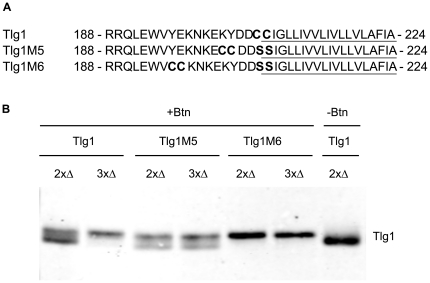
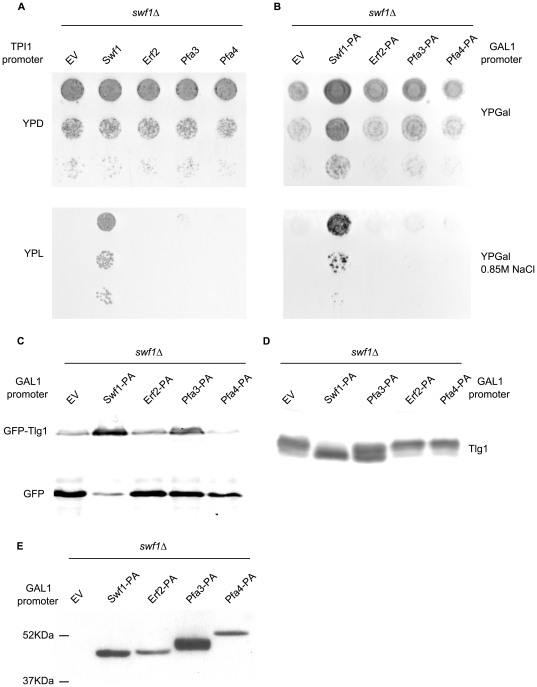
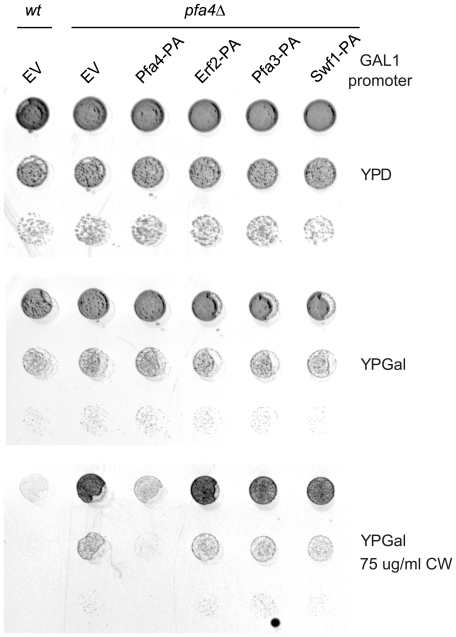
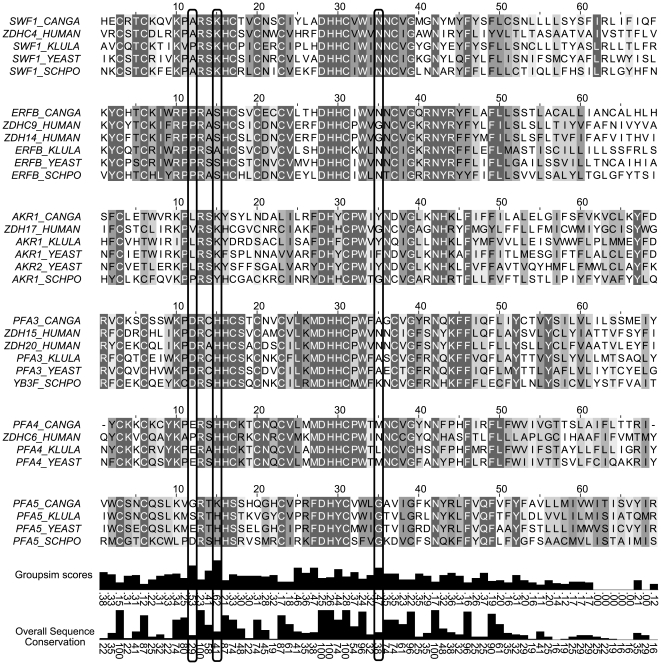
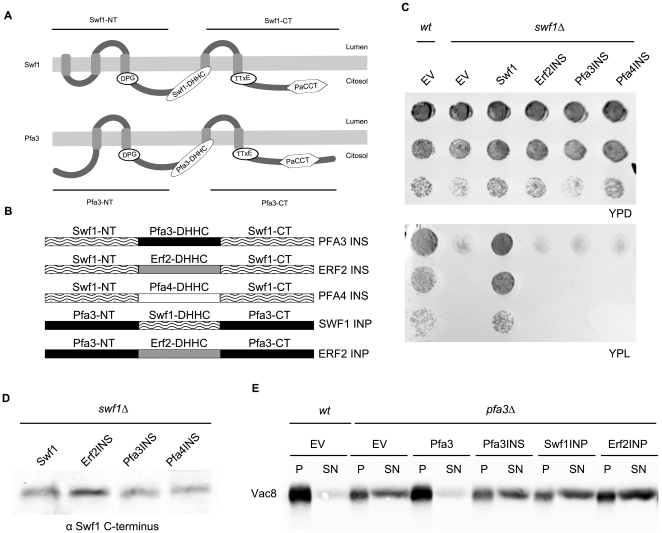
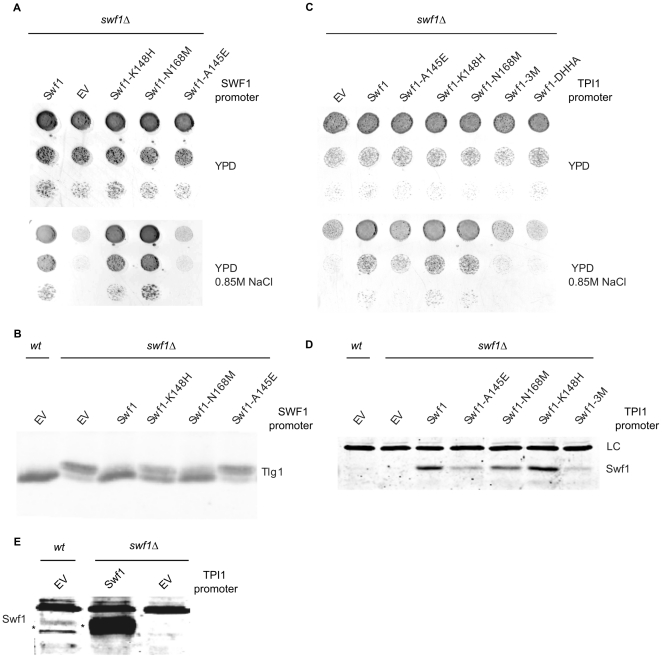
Many proteins are modified after their synthesis, by the addition of a lipid molecule to one or more cysteine residues, through a thioester bond. This modification is called S-acylation, and more commonly palmitoylation. This reaction is carried out by a family of enzymes, called palmitoyltransferases (PATs), characterized by the presence of a conserved 50- aminoacids domain called "Asp-His-His-Cys- Cysteine Rich Domain" (DHHC-CRD). There are 7 members of this family in the yeast Saccharomyces cerevisiae, and each of these proteins is thought to be responsible for the palmitoylation of a subset of substrates. Substrate specificity of PATs, however, is not yet fully understood. Several yeast PATs seem to have overlapping specificity, and it has been proposed that the machinery responsible for palmitoylating peripheral membrane proteins in mammalian cells, lacks specificity altogether.Here we investigate the specificity of transmembrane protein palmitoylation in S. cerevisiae, which is carried out predominantly by two PATs, Swf1 and Pfa4. We show that palmitoylation of transmembrane substrates requires dedicated PATs, since other yeast PATs are mostly unable to perform Swf1 or Pfa4 functions, even when overexpressed. Furthermore, we find that Swf1 is highly specific for its substrates, as it is unable to substitute for other PATs. To identify where Swf1 specificity lies, we carried out a bioinformatics survey to identify amino acids responsible for the determination of specificity or Specificity Determination Positions (SDPs) and showed experimentally, that mutation of the two best SDP candidates, A145 and K148, results in complete and partial loss of function, respectively. These residues are located within the conserved catalytic DHHC domain suggesting that it could also be involved in the determination of specificity. Finally, we show that modifying the position of the cysteines in Tlg1, a Swf1 substrate, results in lack of palmitoylation, as expected for a highly specific enzymatic reaction.
许多蛋白质在合成后会通过硫酯键被添加一个脂质分子到一个或多个半胱氨酸残基上来进行修饰。这种修饰称为 S-酰化,更常见的是棕榈酰化。这个反应是由一类称为棕榈酰转移酶 (PAT) 的酶来完成的,这些酶的特征是存在一个保守的 50 个氨基酸的结构域,称为“天冬氨酸-组氨酸-组氨酸-半胱氨酸丰富结构域”(DHHC-CRD)。在酵母酿酒酵母中有 7 个这样的家族成员,每个成员都被认为负责一组底物的棕榈酰化。然而,PAT 的底物特异性尚未完全理解。一些酵母 PAT 似乎具有重叠的特异性,并且有人提出负责棕榈酰化哺乳动物细胞中周边膜蛋白的机制完全没有特异性。在这里,我们研究了 S. cerevisiae 中跨膜蛋白棕榈酰化的特异性,这主要由两个 PAT,Swf1 和 Pfa4 来完成。我们表明,跨膜底物的棕榈酰化需要专门的 PAT,因为其他酵母 PAT 大多无法执行 Swf1 或 Pfa4 的功能,即使过度表达也是如此。此外,我们发现 Swf1 对其底物具有高度特异性,因为它无法替代其他 PAT。为了确定 Swf1 的特异性所在,我们进行了生物信息学调查以确定决定特异性的氨基酸或特异性决定位置 (SDP),并通过实验表明,突变两个最佳 SDP 候选者 A145 和 K148 分别导致完全和部分功能丧失。这些残基位于保守的催化 DHHC 结构域内,这表明它也可能参与特异性的决定。最后,我们表明,修饰 Tlg1(Swf1 的底物)中半胱氨酸的位置会导致缺乏棕榈酰化,这与高度特异性的酶反应相符。