Department of Biochemistry and Molecular Biology, University of Chicago, Chicago, IL 60637, USA.
J Gen Physiol. 2012 Nov;140(5):481-93. doi: 10.1085/jgp.201210817. Epub 2012 Oct 15.
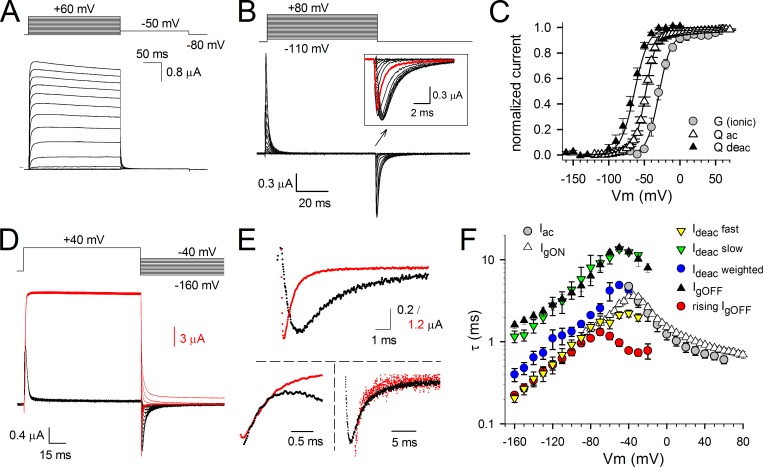
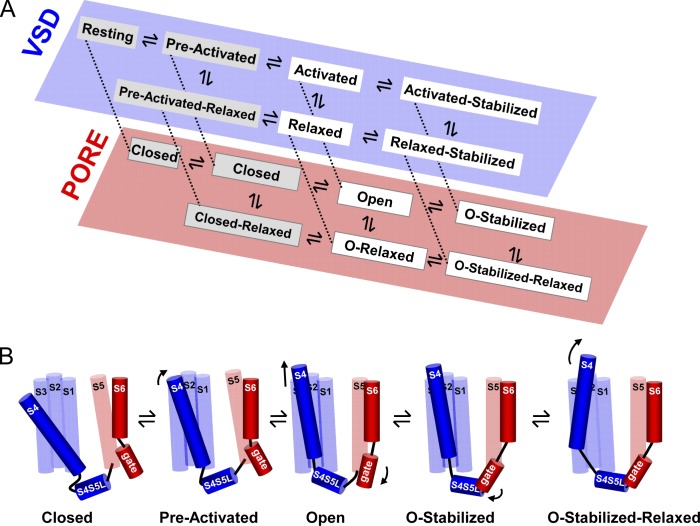
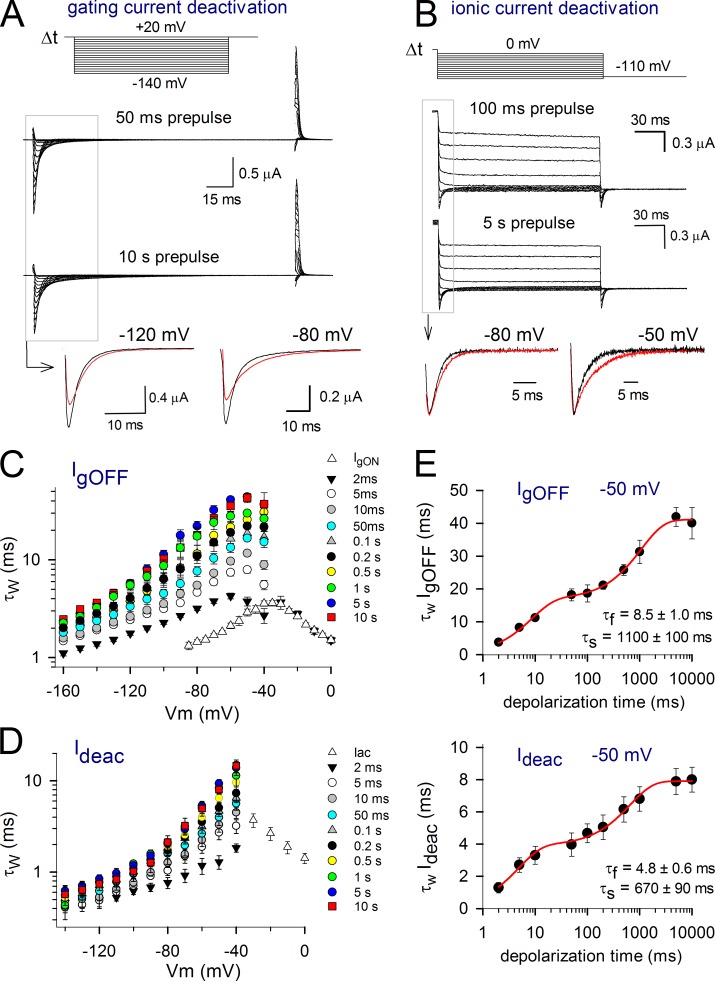
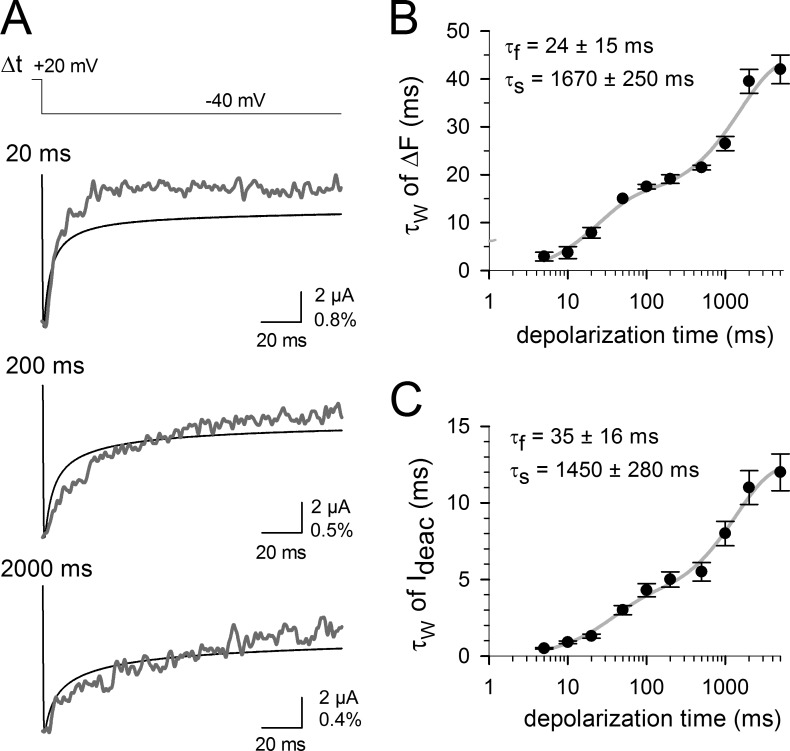
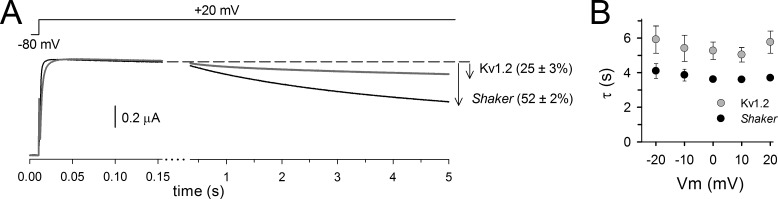
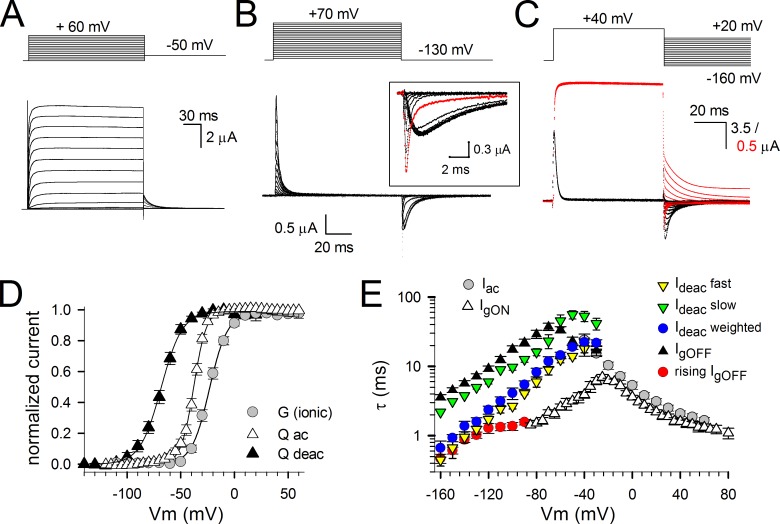
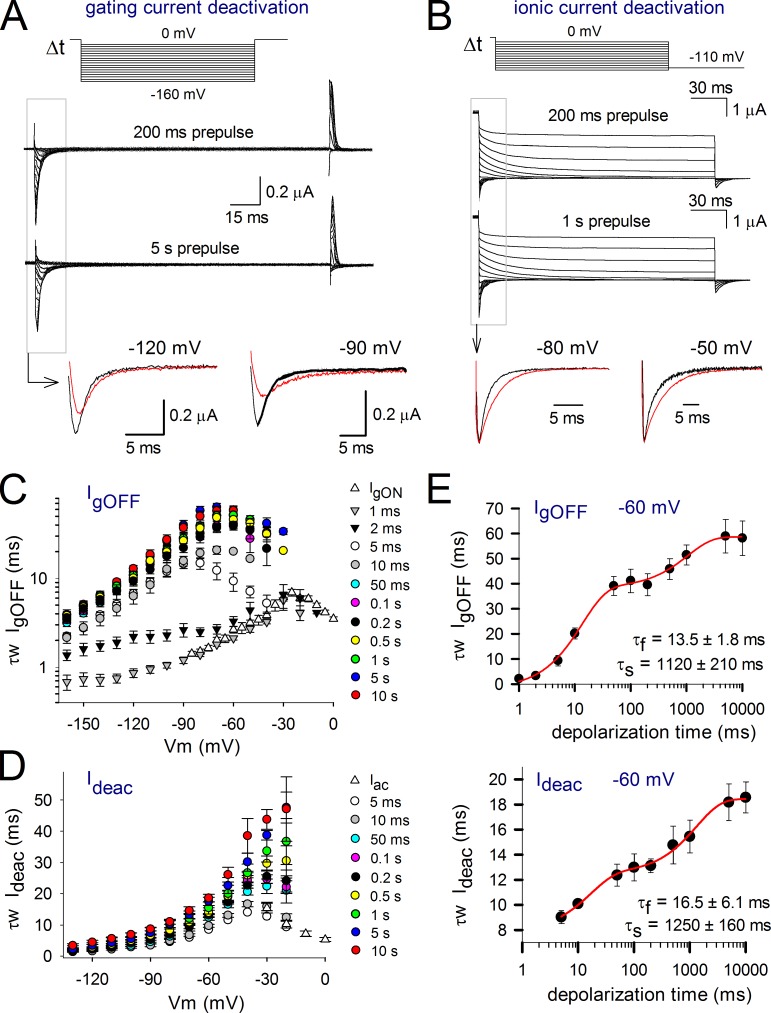
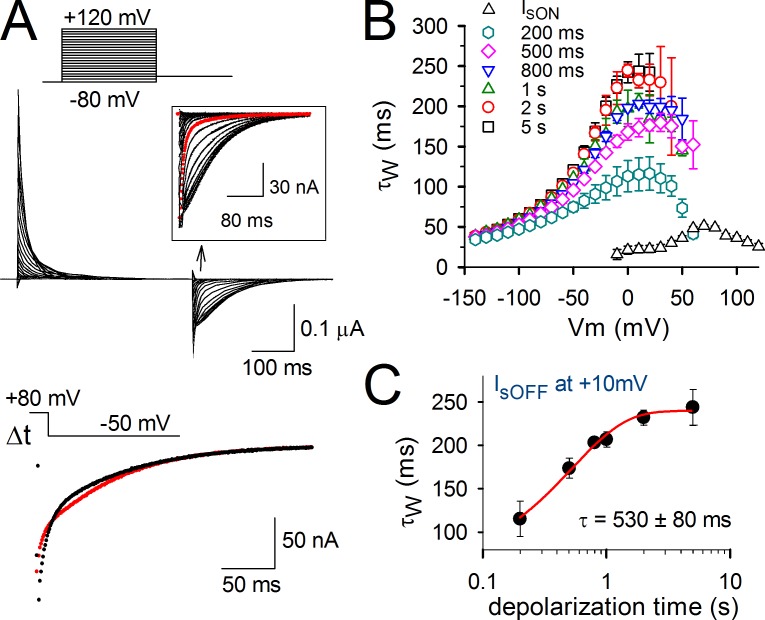
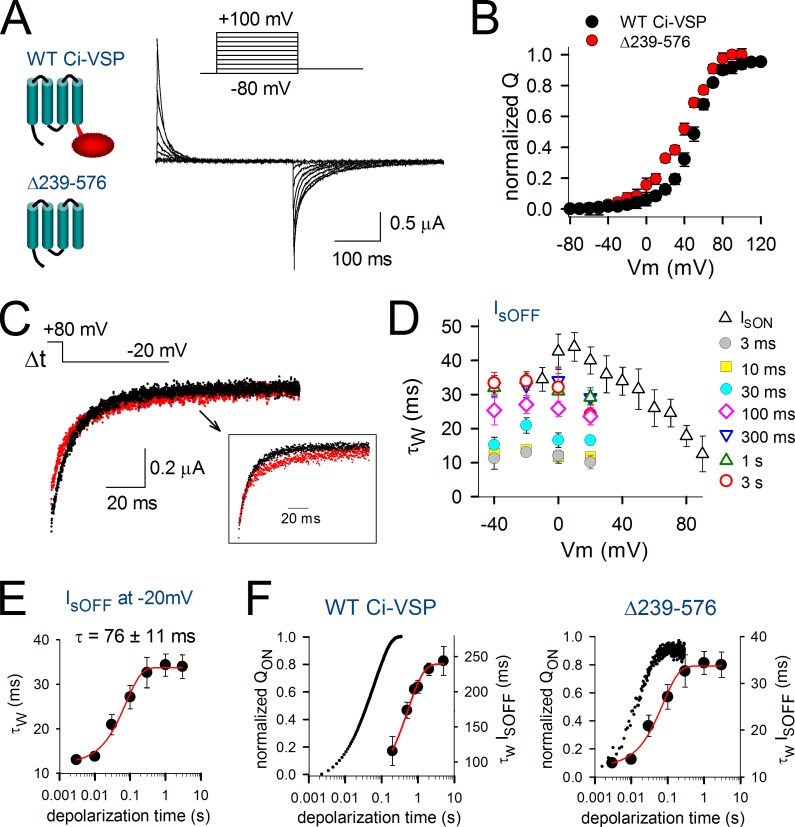
Voltage-dependent potassium (Kv) channels provide the repolarizing power that shapes the action potential duration and helps control the firing frequency of neurons. The K(+) permeation through the channel pore is controlled by an intracellularly located bundle-crossing (BC) gate that communicates with the voltage-sensing domains (VSDs). During prolonged membrane depolarizations, most Kv channels display C-type inactivation that halts K(+) conduction through constriction of the K(+) selectivity filter. Besides triggering C-type inactivation, we show that in Shaker and Kv1.2 channels (expressed in Xenopus laevis oocytes), prolonged membrane depolarizations also slow down the kinetics of VSD deactivation and BC gate closure during the subsequent membrane repolarization. Measurements of deactivating gating currents (reporting VSD movement) and ionic currents (BC gate status) showed that the kinetics of both slowed down in two distinct phases with increasing duration of the depolarizing prepulse. The biphasic slowing in VSD deactivation and BC gate closure was strongly correlated in time and magnitude. Simultaneous recordings of ionic currents and fluorescence from a probe tracking VSD movement in Shaker directly demonstrated that both processes were synchronized. Whereas the first slowing originates from a stabilization imposed by BC gate opening, the subsequent slowing reflects the rearrangement of the VSD toward its relaxed state (relaxation). The VSD relaxation was observed in the Ciona intestinalis voltage-sensitive phosphatase and in its isolated VSD. Collectively, our results show that the VSD relaxation is not kinetically related to C-type inactivation and is an intrinsic property of the VSD. We propose VSD relaxation as a general mechanism for depolarization-induced slowing of BC gate closure that may enable Kv1.2 channels to modulate the firing frequency of neurons based on the depolarization history.
电压门控钾 (Kv) 通道提供了复极化能力,这决定了动作电位的持续时间,并有助于控制神经元的放电频率。通道孔内的 K(+) 通过位于细胞内的束交叉 (BC) 门控来控制,BC 门控与电压感应域 (VSD) 通讯。在长时间的膜去极化期间,大多数 Kv 通道表现出 C 型失活,通过 K(+) 选择性滤器的收缩来阻止 K(+) 传导。除了触发 C 型失活外,我们还表明,在 Shaker 和 Kv1.2 通道(在非洲爪蟾卵母细胞中表达)中,长时间的膜去极化也会在随后的膜复极化期间减缓 VSD 去激活和 BC 门控关闭的动力学。失活动力学电流(报告 VSD 运动)和离子电流(BC 门控状态)的测量表明,随着去极化预脉冲持续时间的增加,两种动力学都以两个不同的相缓慢下来。VSD 去激活和 BC 门控关闭的双相减速在时间和幅度上都有很强的相关性。Shaker 中离子电流和荧光同时记录,直接证明了这两个过程是同步的。虽然第一个减速源于 BC 门控打开所引起的稳定化,但随后的减速反映了 VSD 向其松弛状态(松弛)的重新排列。在海鞘电压敏感磷酸酶及其分离的 VSD 中观察到 VSD 松弛。总的来说,我们的结果表明,VSD 松弛在动力学上与 C 型失活无关,是 VSD 的固有特性。我们提出 VSD 松弛是一种普遍的机制,用于解释去极化诱导的 BC 门控关闭减速,这可能使 Kv1.2 通道能够根据去极化历史来调节神经元的放电频率。