Department of Cell Biology, Microbiology, and Molecular Biology, University of South Florida Tampa, Florida, USA.
PLoS One. 2013;8(3):e58345. doi: 10.1371/journal.pone.0058345. Epub 2013 Mar 5.
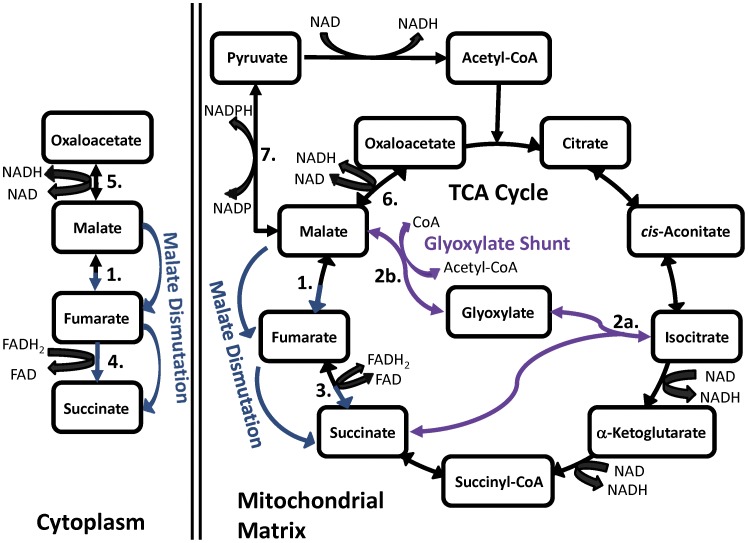
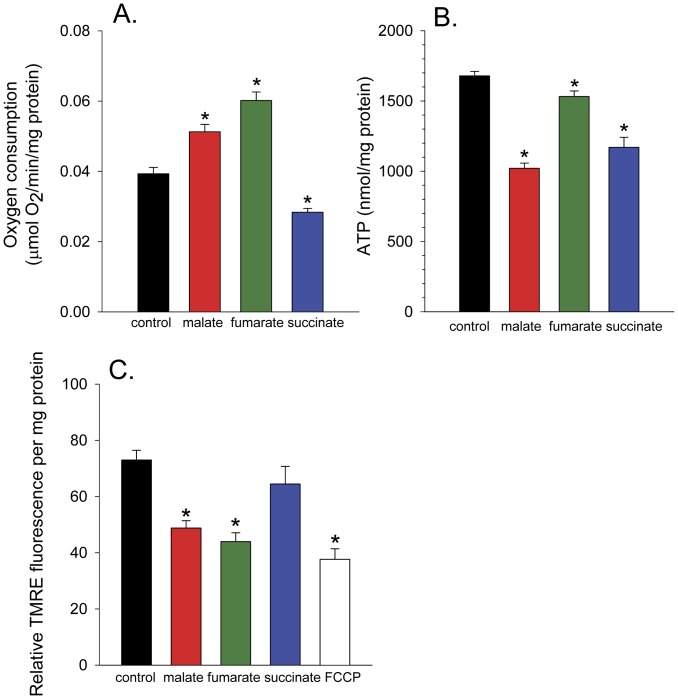
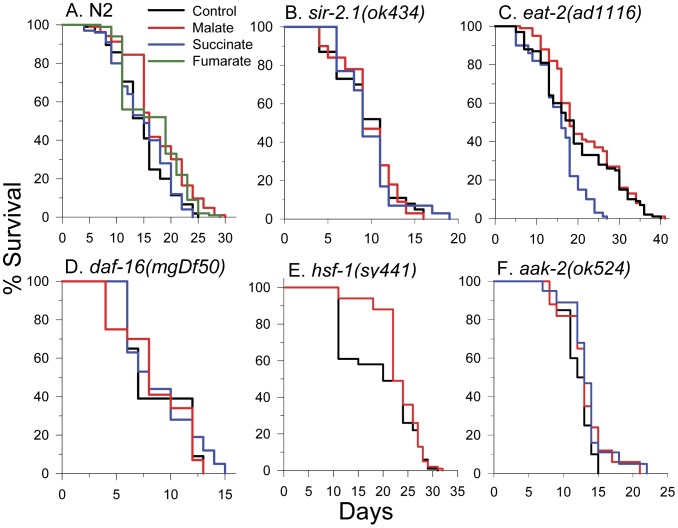
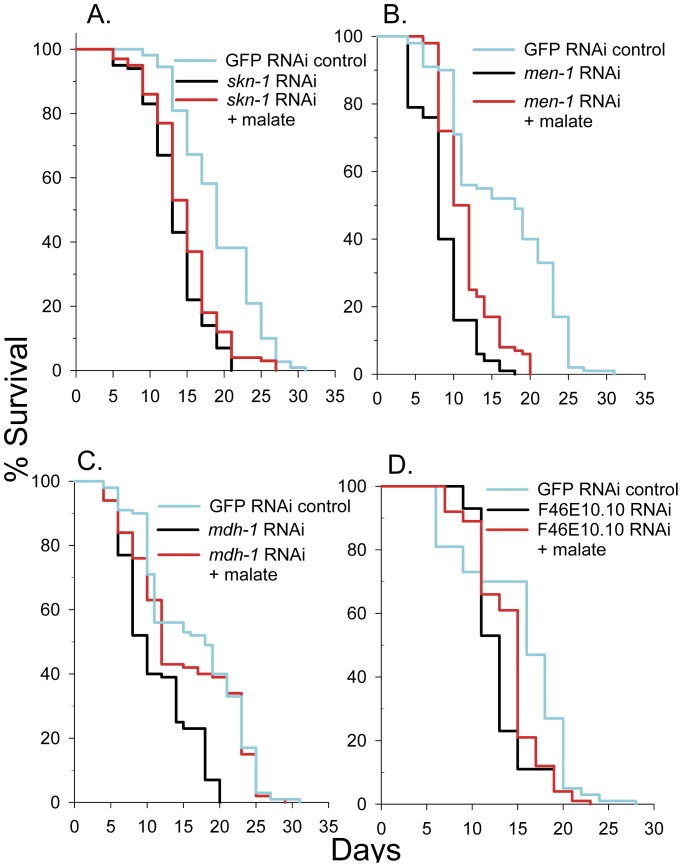
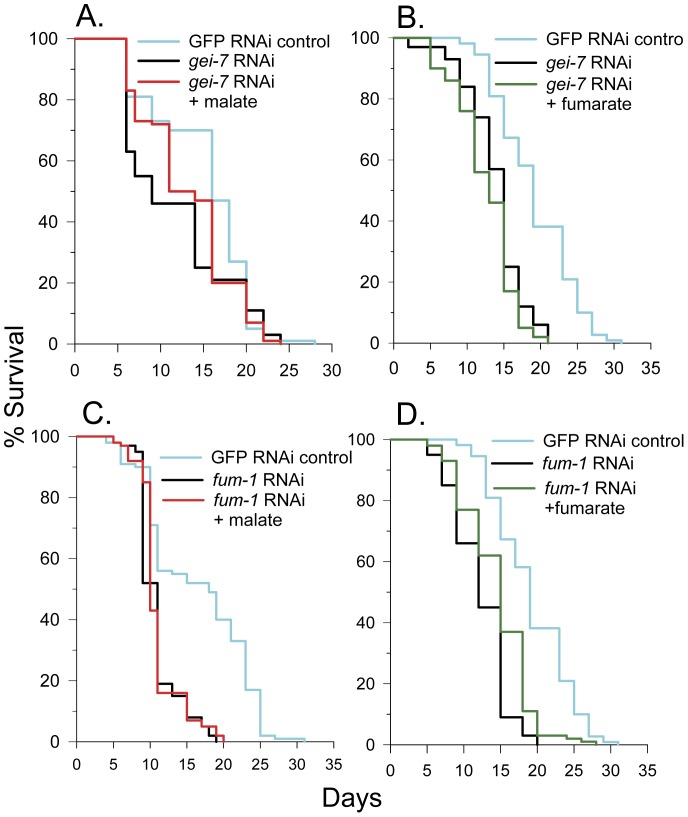
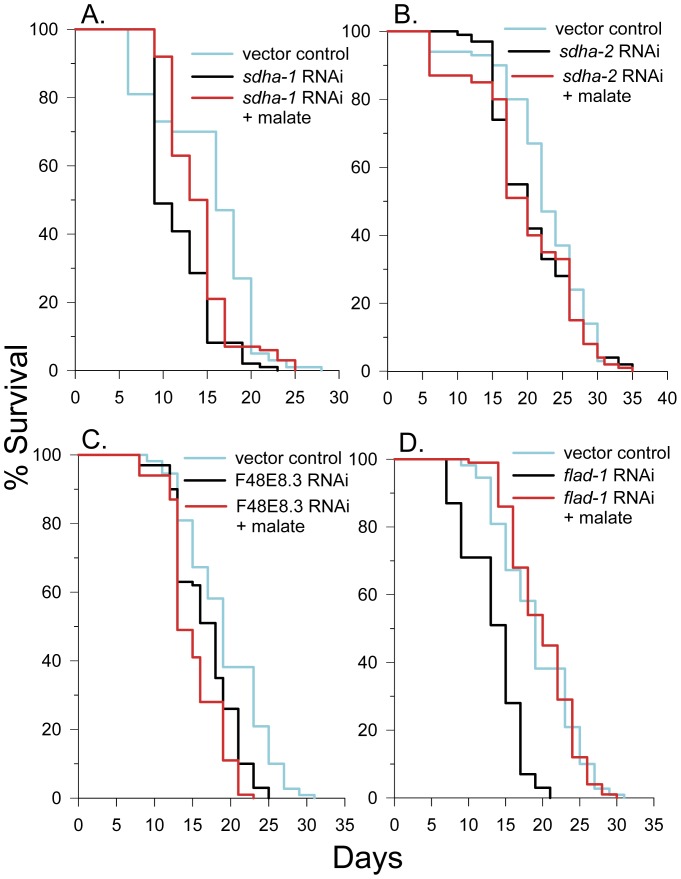
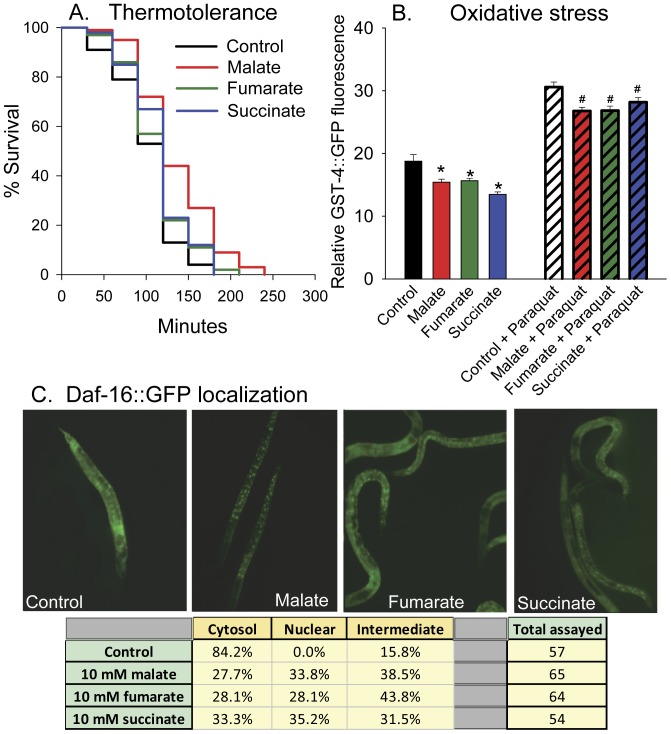
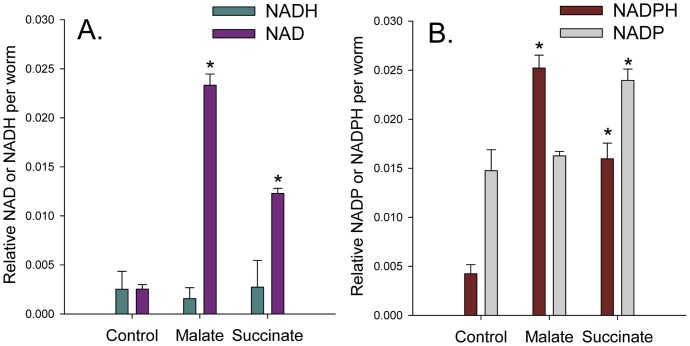
Malate, the tricarboxylic acid (TCA) cycle metabolite, increased lifespan and thermotolerance in the nematode C. elegans. Malate can be synthesized from fumarate by the enzyme fumarase and further oxidized to oxaloacetate by malate dehydrogenase with the accompanying reduction of NAD. Addition of fumarate also extended lifespan, but succinate addition did not, although all three intermediates activated nuclear translocation of the cytoprotective DAF-16/FOXO transcription factor and protected from paraquat-induced oxidative stress. The glyoxylate shunt, an anabolic pathway linked to lifespan extension in C. elegans, reversibly converts isocitrate and acetyl-CoA to succinate, malate, and CoA. The increased longevity provided by malate addition did not occur in fumarase (fum-1), glyoxylate shunt (gei-7), succinate dehydrogenase flavoprotein (sdha-2), or soluble fumarate reductase F48E8.3 RNAi knockdown worms. Therefore, to increase lifespan, malate must be first converted to fumarate, then fumarate must be reduced to succinate by soluble fumarate reductase and the mitochondrial electron transport chain complex II. Reduction of fumarate to succinate is coupled with the oxidation of FADH2 to FAD. Lifespan extension induced by malate depended upon the longevity regulators DAF-16 and SIR-2.1. Malate supplementation did not extend the lifespan of long-lived eat-2 mutant worms, a model of dietary restriction. Malate and fumarate addition increased oxygen consumption, but decreased ATP levels and mitochondrial membrane potential suggesting a mild uncoupling of oxidative phosphorylation. Malate also increased NADPH, NAD, and the NAD/NADH ratio. Fumarate reduction, glyoxylate shunt activity, and mild mitochondrial uncoupling likely contribute to the lifespan extension induced by malate and fumarate by increasing the amount of oxidized NAD and FAD cofactors.
苹果酸是三羧酸(TCA)循环代谢物,可延长线虫 C. elegans 的寿命和耐热性。苹果酸可由酶延胡索酸酶从富马酸合成,然后进一步被苹果酸脱氢酶氧化为草酰乙酸,同时伴随着 NAD 的还原。添加富马酸也延长了寿命,但添加琥珀酸则没有,尽管这三种中间产物都激活了细胞保护因子 DAF-16/FOXO 的核转位,并防止百草枯诱导的氧化应激。乙醛酸支路是一种与线虫寿命延长相关的合成代谢途径,可将异柠檬酸和乙酰辅酶 A 可逆地转化为琥珀酸、苹果酸和辅酶 A。添加苹果酸所提供的长寿作用不会发生在延胡索酸酶(fum-1)、乙醛酸支路(gei-7)、琥珀酸脱氢酶黄素蛋白(sdha-2)或可溶性延胡索酸还原酶 F48E8.3 RNAi 敲低线虫中。因此,要延长寿命,苹果酸必须首先转化为富马酸,然后可溶性延胡索酸还原酶和线粒体电子传递链复合物 II 将富马酸还原为琥珀酸。富马酸还原为琥珀酸与 FADH2 氧化为 FAD 偶联。苹果酸诱导的寿命延长取决于长寿调节剂 DAF-16 和 SIR-2.1。苹果酸补充剂不能延长长寿的 eat-2 突变体线虫的寿命,这是一种饮食限制的模型。苹果酸和富马酸的添加增加了耗氧量,但降低了 ATP 水平和线粒体膜电位,表明氧化磷酸化轻度解偶联。苹果酸还增加了 NADPH、NAD 和 NAD/NADH 比值。富马酸还原、乙醛酸支路活性和轻度线粒体解偶联可能通过增加氧化的 NAD 和 FAD 辅因子的量,有助于苹果酸和富马酸诱导的寿命延长。