Department of Chemistry, The Pennsylvania State University, University Park, PA 16802, USA.
Biochemistry. 2013 Apr 30;52(17):2874-87. doi: 10.1021/bi400136u. Epub 2013 Apr 16.
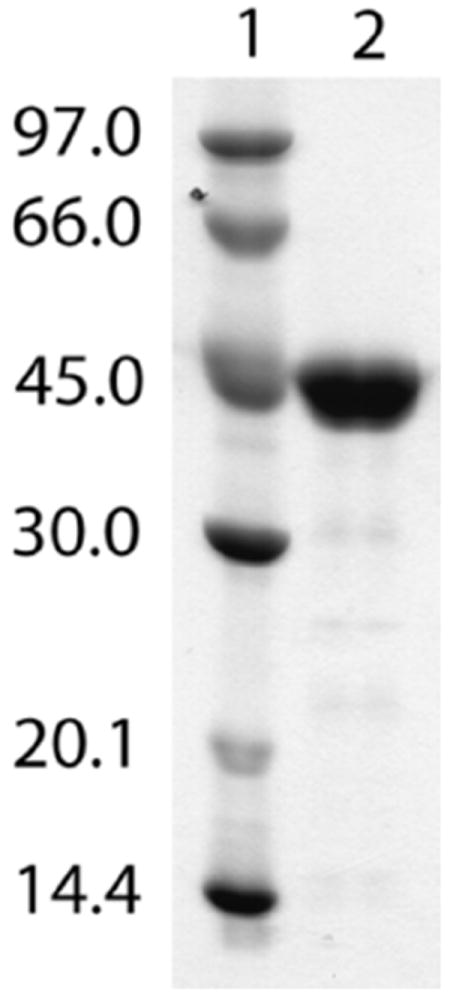
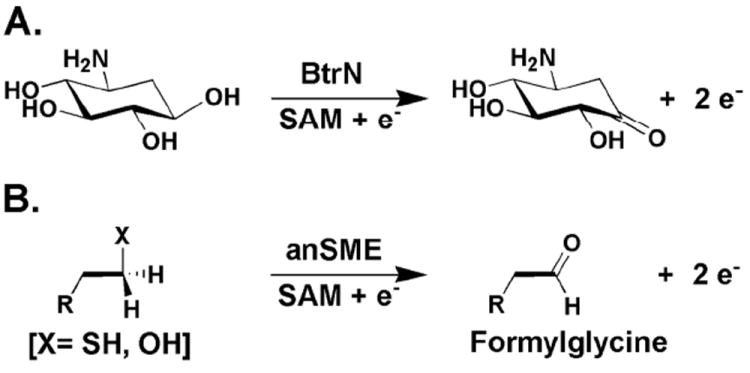
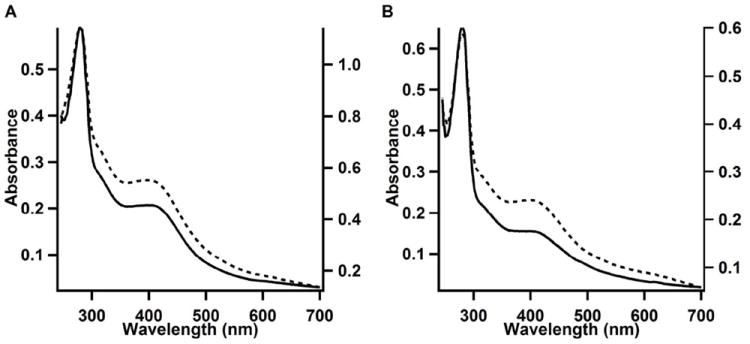
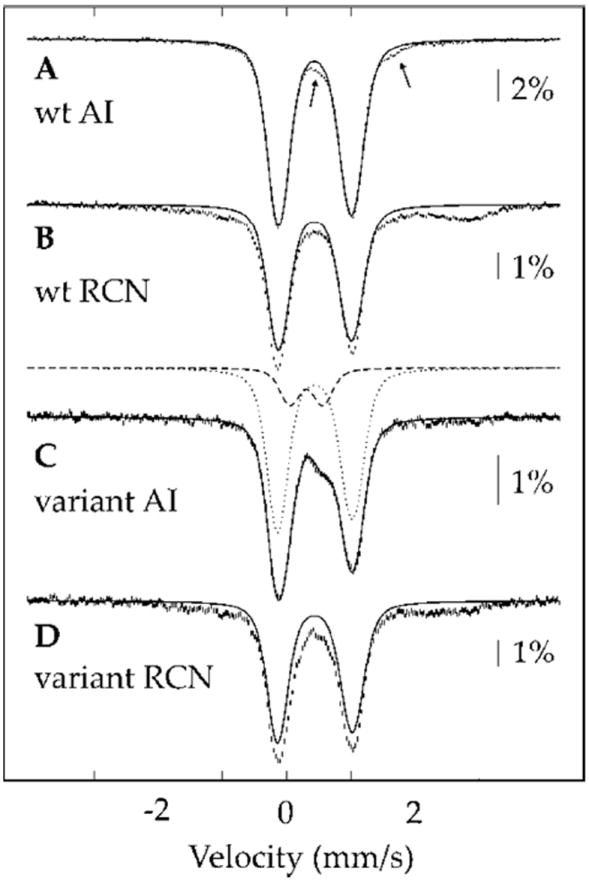
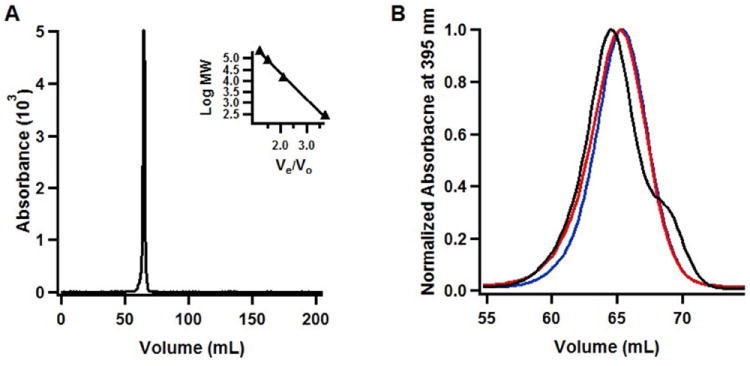
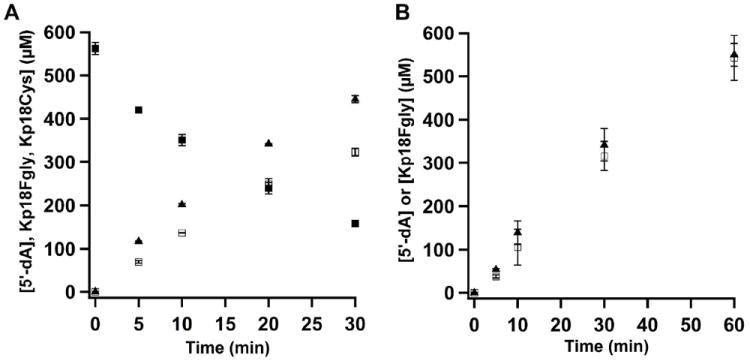
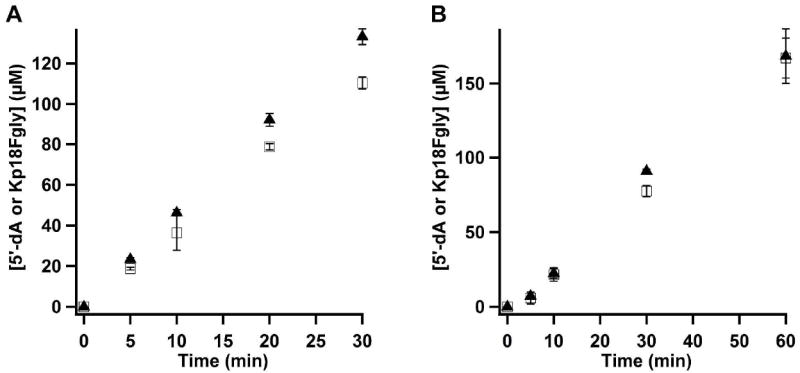
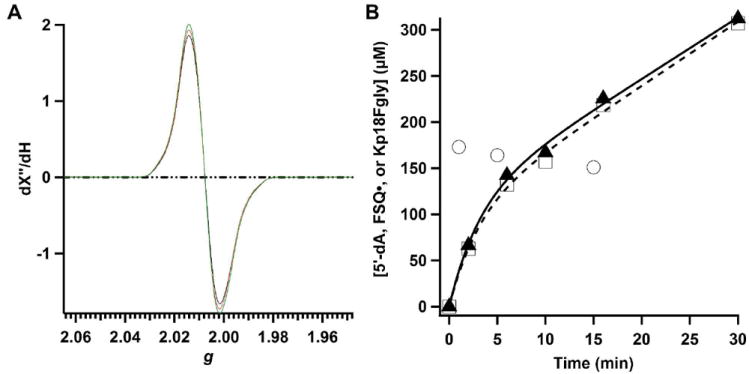
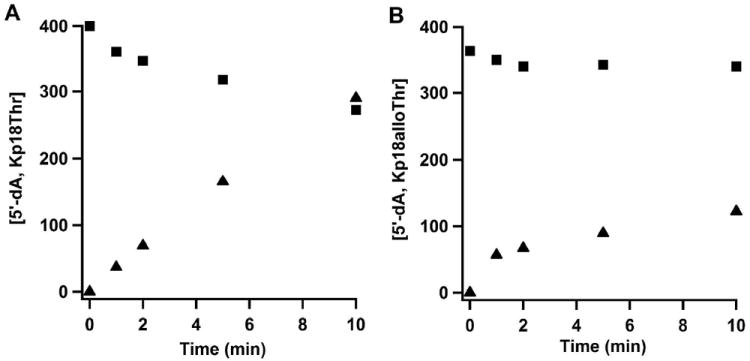
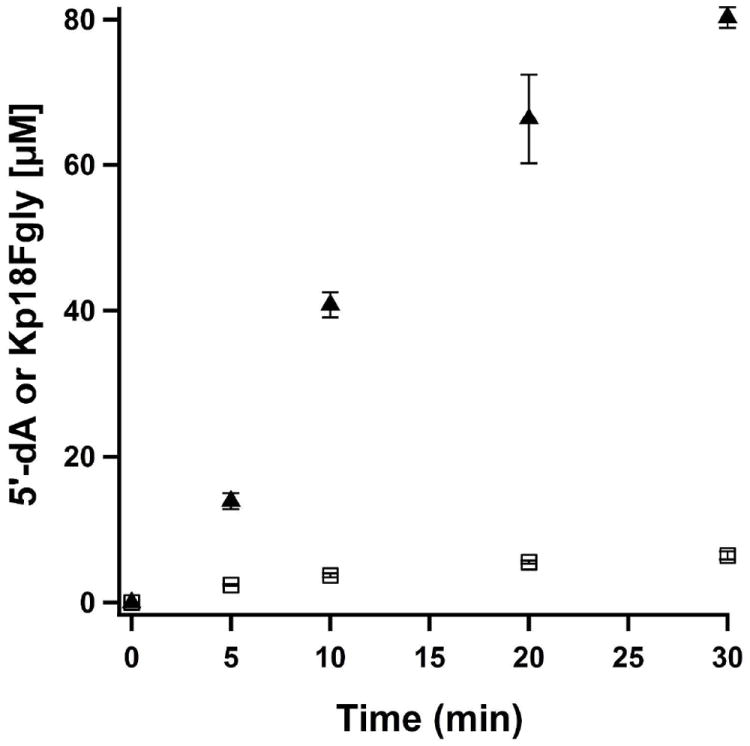
The anaerobic sulfatase-maturating enzyme from Clostridium perfringens (anSMEcpe) catalyzes the two-electron oxidation of a cysteinyl residue on a cognate protein to a formylglycyl residue (FGly) using a mechanism that involves organic radicals. The FGly residue plays a unique role as a cofactor in a class of enzymes termed arylsulfatases, which catalyze the hydrolysis of various organosulfate monoesters. anSMEcpe has been shown to be a member of the radical S-adenosylmethionine (SAM) family of enzymes, [4Fe-4S] cluster-requiring proteins that use a 5'-deoxyadenosyl 5'-radical (5'-dA(•)) generated from a reductive cleavage of SAM to initiate radical-based catalysis. Herein, we show that anSMEcpe contains in addition to the [4Fe-4S] cluster harbored by all radical SAM (RS) enzymes, two additional [4Fe-4S] clusters, similar to the radical SAM protein AtsB, which catalyzes the two-electron oxidation of a seryl residue to a FGly residue. We show by size-exclusion chromatography that both AtsB and anSMEcpe are monomeric proteins, and site-directed mutagenesis studies of AtsB reveal that individual Cys → Ala substitutions at seven conserved positions result in an insoluble protein, consistent with those residues acting as ligands to the two additional [4Fe-4S] clusters. Ala substitutions at an additional conserved Cys residue (C291 in AtsB and C276 in anSMEcpe) afford proteins that display intermediate behavior. These proteins exhibit reduced solubility and drastically reduced activity, behavior that is conspicuously similar to that of a critical Cys residue in BtrN, another radical SAM dehydrogenase [Grove, T. L., et al. (2010) Biochemistry 49, 3783-3785]. We also show that wild-type anSMEcpe acts on peptides containing other oxidizable amino acids at the target position. Moreover, we show that the enzyme will convert threonyl peptides to the corresponding ketone product, and also allo-threonyl peptides, but with a significantly reduced efficiency, suggesting that the pro-S hydrogen atom of the normal cysteinyl substrate is stereoselectively removed during turnover. Lastly, we show that the electron generated during catalysis by AtsB and anSMEcpe can be utilized for multiple turnovers, albeit through a reduced flavodoxin-mediated pathway.
产气荚膜梭菌的厌氧磺基转移酶成熟酶(anSMEcpe)使用涉及有机自由基的机制,催化同源蛋白上半胱氨酸残基的两电子氧化为甲酰甘氨酸残基(FGly)。FGly 残基作为一类称为芳基硫酸酯酶的酶的辅因子发挥独特作用,芳基硫酸酯酶催化各种有机硫酸单酯的水解。anSMEcpe 已被证明是自由基 S-腺苷甲硫氨酸(SAM)家族酶的成员,[4Fe-4S]簇需要的蛋白质,使用 SAM 的还原裂解产生的 5'-脱氧腺苷 5'-自由基(5'-dA(•))来启动基于自由基的催化。在此,我们表明,anSMEcpe 除了所有自由基 SAM(RS)酶所具有的 [4Fe-4S]簇之外,还含有两个额外的 [4Fe-4S]簇,类似于催化丝氨酸残基两电子氧化为 FGly 残基的自由基 SAM 蛋白 AtsB。通过凝胶过滤色谱法表明,AtsB 和 anSMEcpe 均为单体蛋白,AtsB 的定点突变研究表明,七个保守位置的单个 Cys→Ala 取代导致不溶性蛋白,这与这些残基作为两个额外的 [4Fe-4S]簇的配体一致。在另外一个保守 Cys 残基(AtsB 中的 C291 和 anSMEcpe 中的 C276)处的 Ala 取代提供了显示中间行为的蛋白质。这些蛋白质表现出降低的溶解度和明显降低的活性,其行为与另一种自由基 SAM 脱氢酶 BtrN 中的关键 Cys 残基明显相似。[Grove, T. L., et al. (2010) Biochemistry 49, 3783-3785]。我们还表明,野生型 anSMEcpe 作用于靶位含有其他可氧化氨基酸的肽。此外,我们表明该酶将将苏氨酸肽转化为相应的酮产物,并且也将 allo-苏氨酸肽转化为相应的酮产物,但效率显著降低,表明在周转过程中,正常半胱氨酸底物的 pro-S 氢原子被立体选择性地去除。最后,我们表明,由 AtsB 和 anSMEcpe 催化产生的电子可以被多次利用,尽管是通过减少的黄素蛋白介导的途径。