Center for Regenerative and Developmental Biology, and Biology Department, Tufts University, Medford, MA 02155, USA.
Dev Biol. 2013 Jul 1;379(1):1-15. doi: 10.1016/j.ydbio.2013.03.021. Epub 2013 Apr 10.
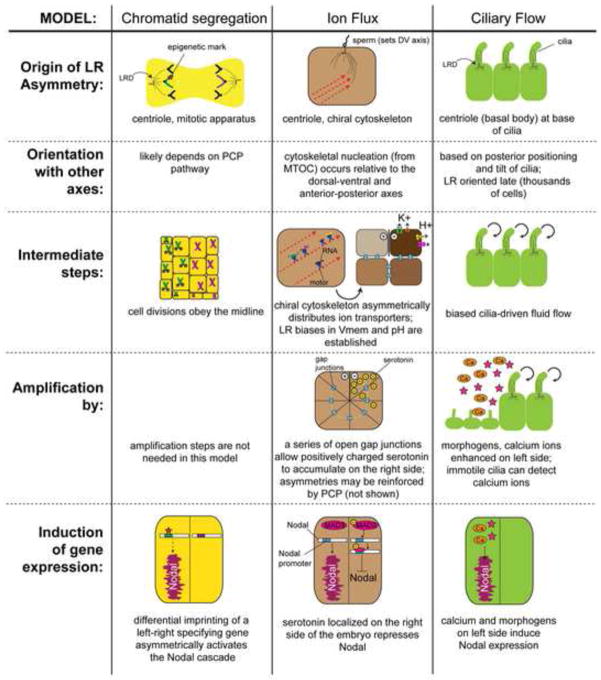
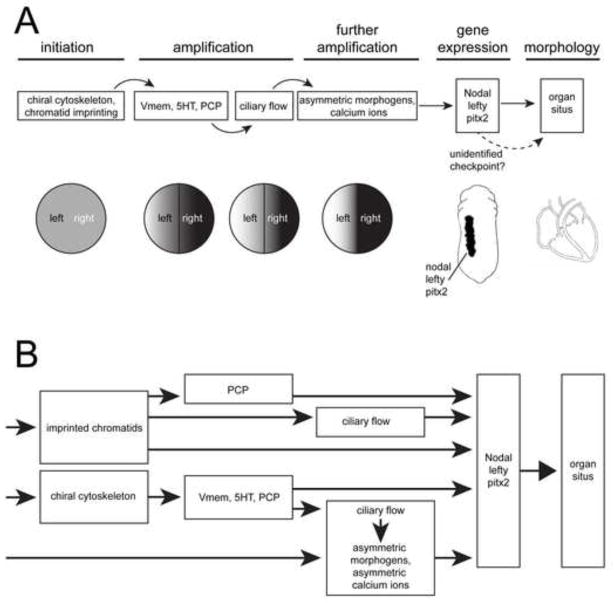
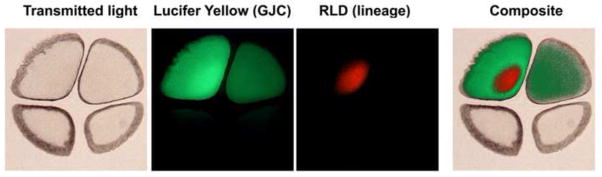
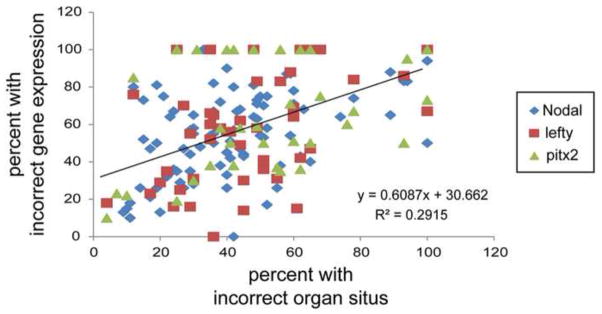
Understanding how and when the left-right (LR) axis is first established is a fundamental question in developmental biology. A popular model is that the LR axis is established relatively late in embryogenesis, due to the movement of motile cilia and the resultant directed fluid flow during late gastrulation/early neurulation. Yet, a large body of evidence suggests that biophysical, molecular, and bioelectrical asymmetries exist much earlier in development, some as early as the first cell cleavage after fertilization. Alternative models of LR asymmetry have been proposed that accommodate these data, postulating that asymmetry is established due to a chiral cytoskeleton and/or the asymmetric segregation of chromatids. There are some similarities, and many differences, in how these various models postulate the origin and timing of symmetry breaking and amplification, and these events' linkage to the well-conserved subsequent asymmetric transcriptional cascades. This review examines experimental data that lend strong support to an early origin of LR asymmetry, yet are also consistent with later roles for cilia in the amplification of LR pathways. In this way, we propose that the various models of asymmetry can be unified: early events are needed to initiate LR asymmetry, and later events could be utilized by some species to maintain LR-biases. We also present an alternative hypothesis, which proposes that individual embryos stochastically choose one of several possible pathways with which to establish their LR axis. These two hypotheses are both tractable in appropriate model species; testing them to resolve open questions in the field of LR patterning will reveal interesting new biology of wide relevance to developmental, cell, and evolutionary biology.
理解左右(LR)轴是如何以及何时首次建立的,是发育生物学中的一个基本问题。一个流行的模型是,由于在原肠胚晚期/神经胚早期运动纤毛的运动和由此产生的定向流体流动,LR 轴在胚胎发生过程中相对较晚建立。然而,大量证据表明,生物物理、分子和生物电学的不对称性在发育过程中很早就存在,有些甚至早在受精后的第一次细胞分裂。已经提出了一些替代的 LR 不对称模型来适应这些数据,假设不对称性是由于手性细胞骨架和/或染色质的不对称分离而建立的。这些各种模型假设对称破缺和放大的起源和时间,以及这些事件与随后保守的不对称转录级联的联系,存在一些相似之处,也存在许多差异。本综述检查了实验数据,这些数据强烈支持 LR 不对称性的早期起源,但也与纤毛在 LR 途径放大中的后期作用一致。通过这种方式,我们提出可以统一各种不对称模型:早期事件需要启动 LR 不对称,而后期事件可以被某些物种用来维持 LR 偏倚。我们还提出了另一种假设,即个体胚胎随机选择几种可能的途径之一来建立它们的 LR 轴。这两种假设在适当的模型物种中都是可行的;通过测试它们来解决 LR 模式领域的悬而未决的问题,将揭示出与发育、细胞和进化生物学广泛相关的有趣的新生物学。