Forest and Wildlife Ecology, University of Wisconsin-Madison, Madison, Wisconsin, USA.
Compr Physiol. 2013 Apr;3(2):741-83. doi: 10.1002/cphy.c110054.
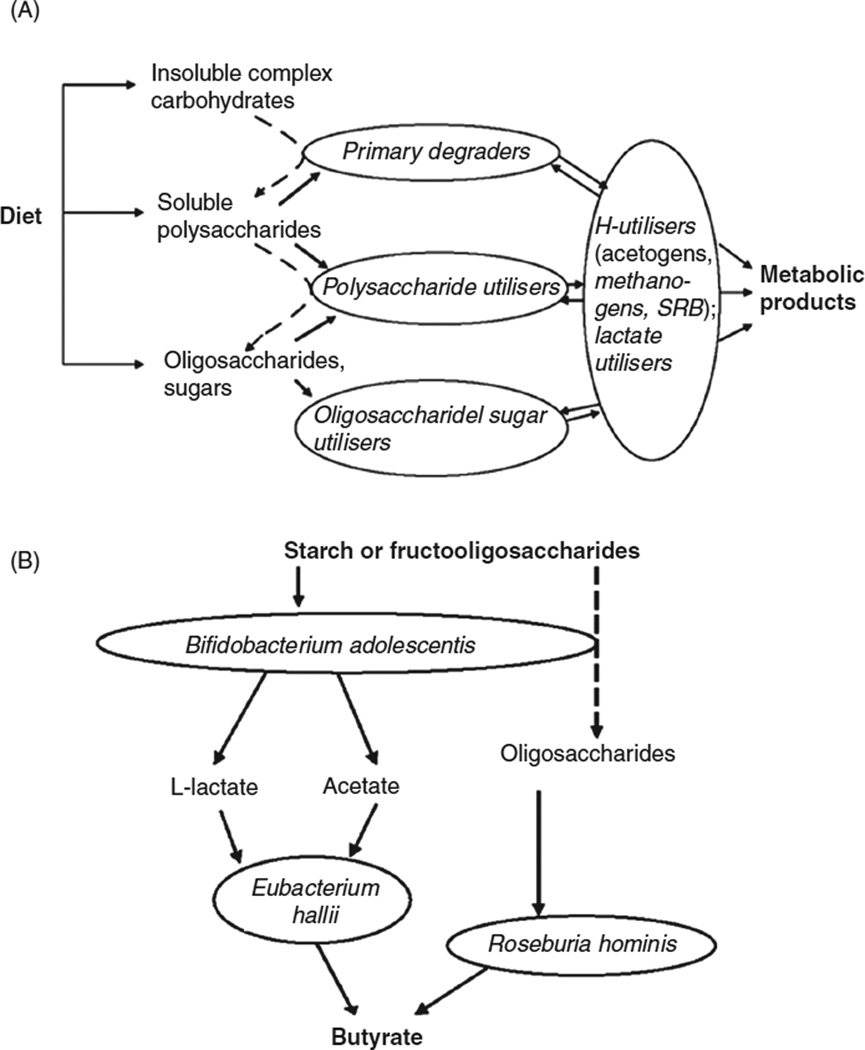
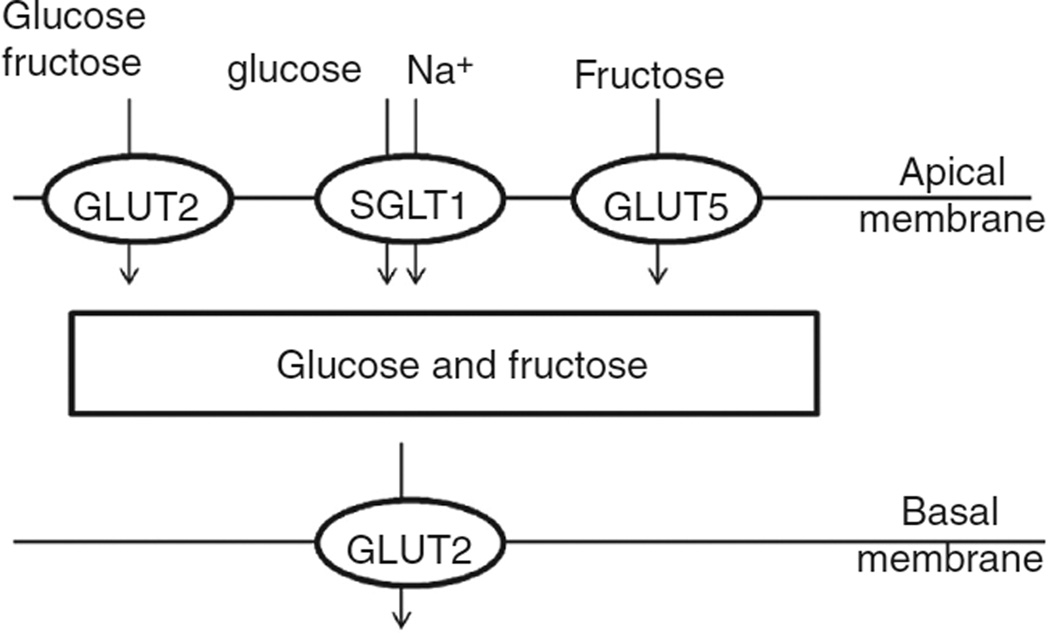
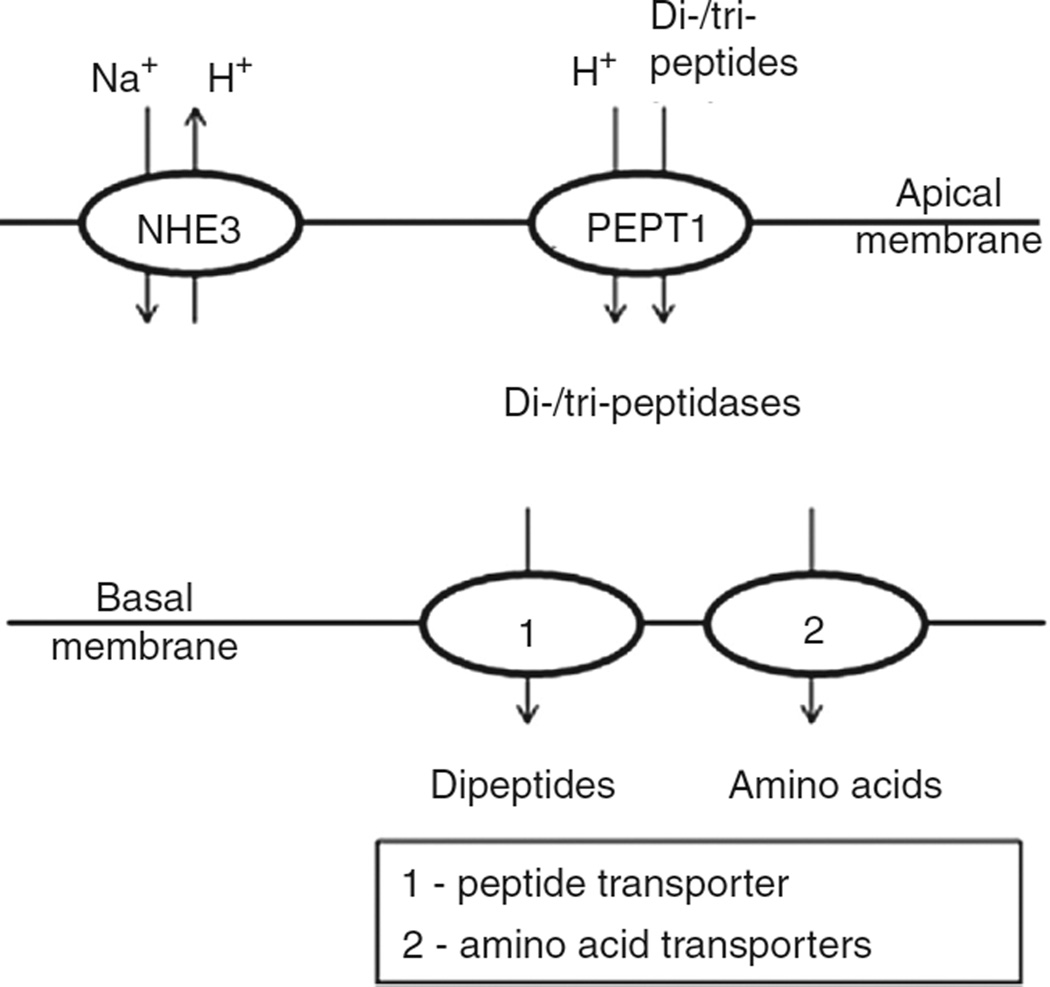
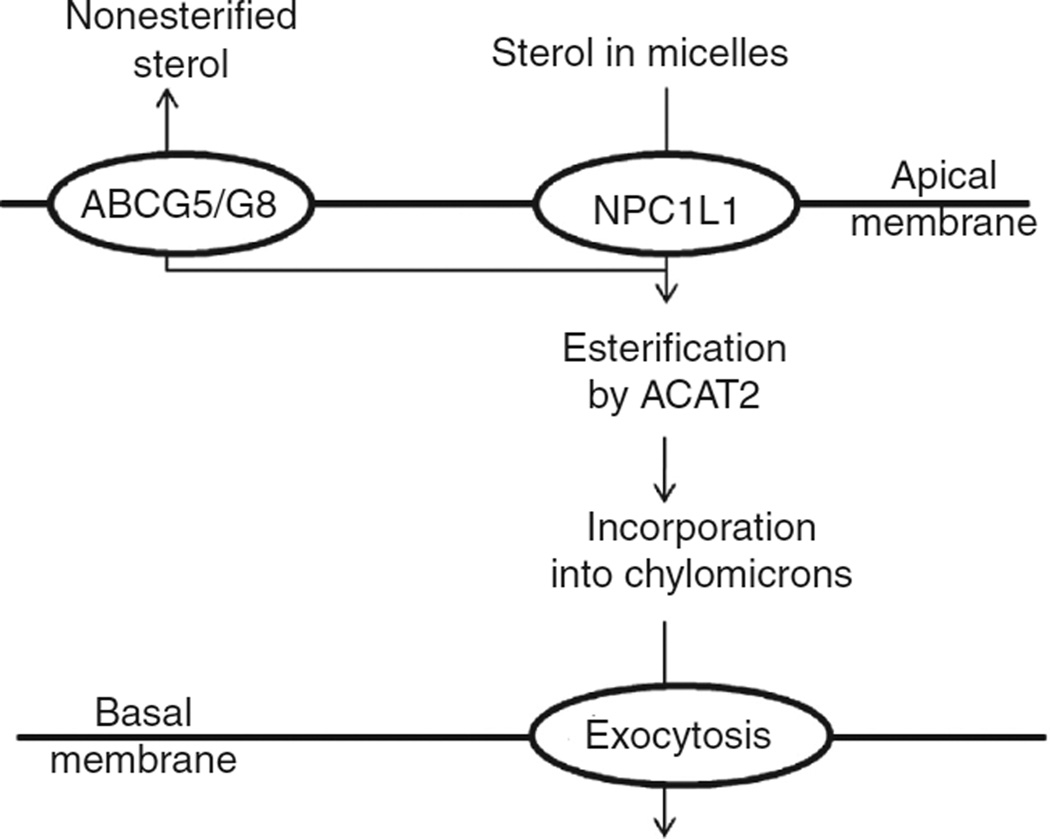
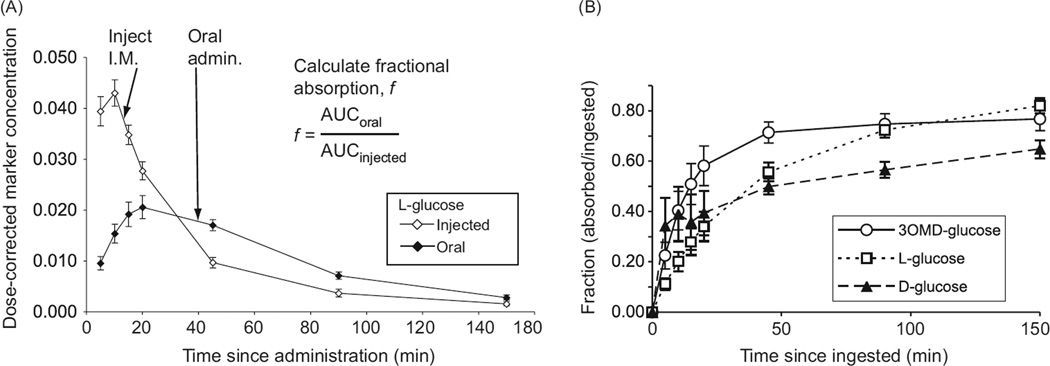
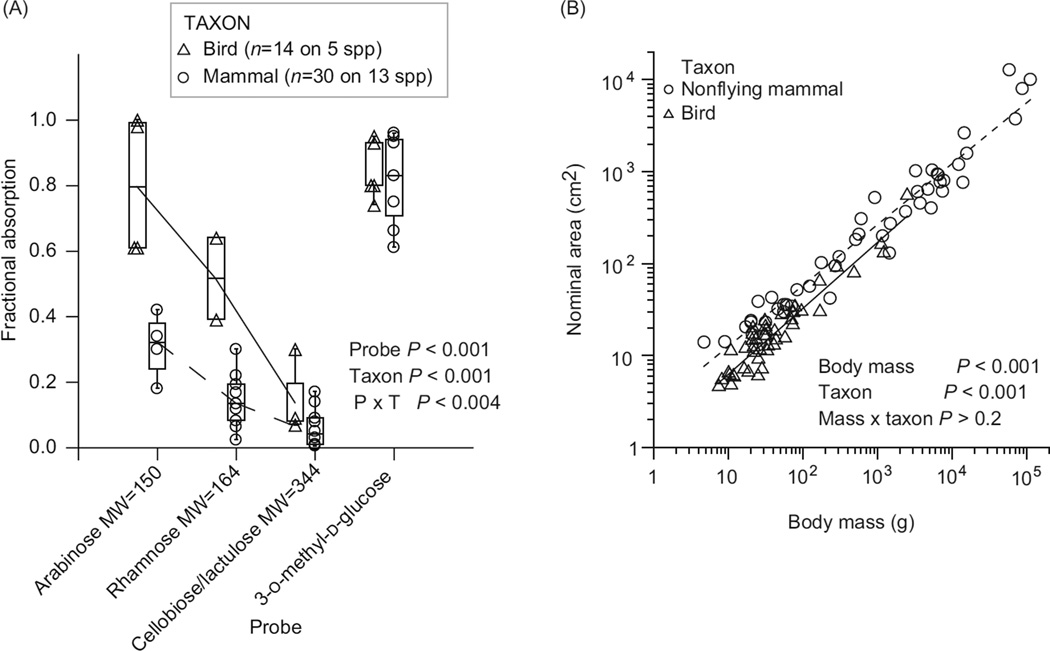
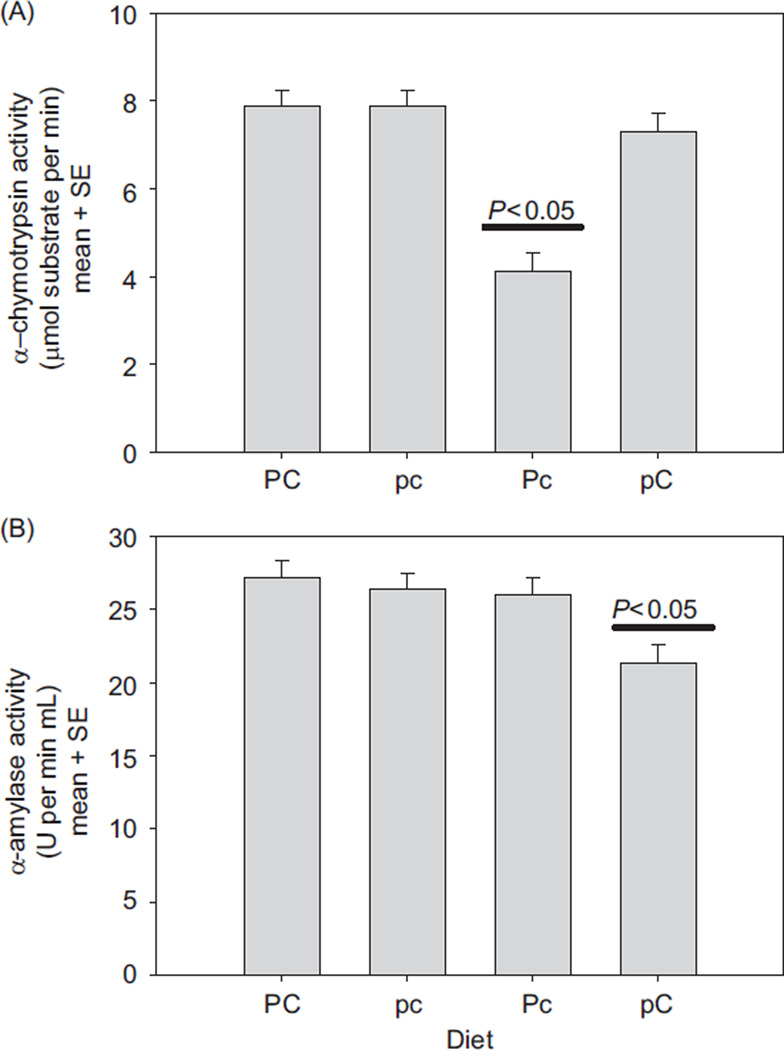
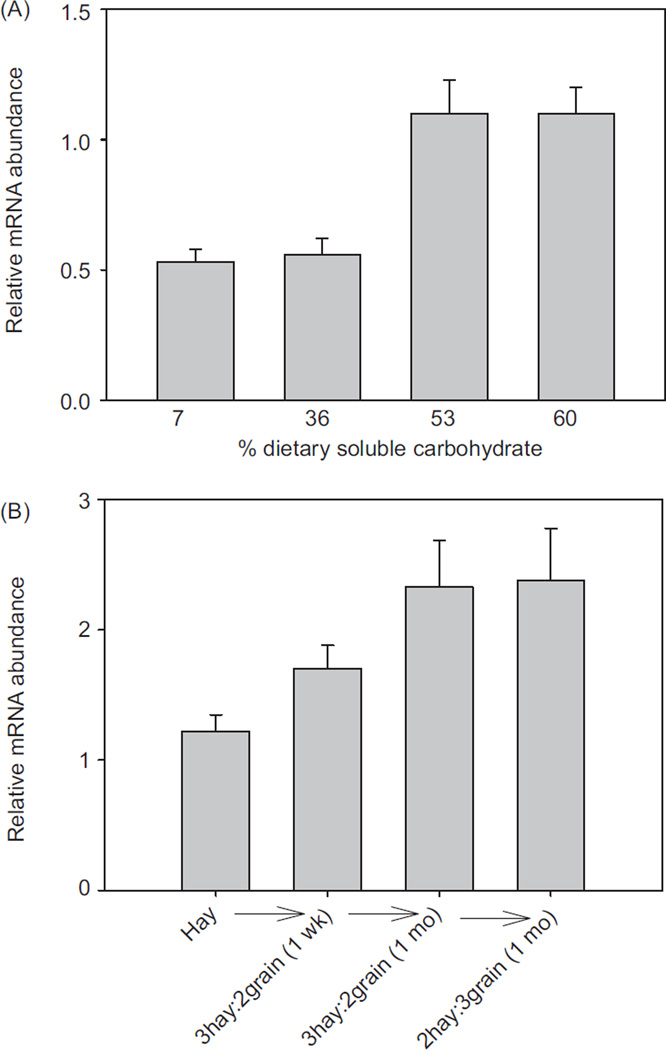
In vertebrates and invertebrates, morphological and functional features of gastrointestinal (GI) tracts generally reflect food chemistry, such as content of carbohydrates, proteins, fats, and material(s) refractory to rapid digestion (e.g., cellulose). The expression of digestive enzymes and nutrient transporters approximately matches the dietary load of their respective substrates, with relatively modest excess capacity. Mechanisms explaining differences in hydrolase activity between populations and species include gene copy number variations and single-nucleotide polymorphisms. Transcriptional and posttranscriptional adjustments mediate phenotypic changes in the expression of hydrolases and transporters in response to dietary signals. Many species respond to higher food intake by flexibly increasing digestive compartment size. Fermentative processes by symbiotic microorganisms are important for cellulose degradation but are relatively slow, so animals that rely on those processes typically possess special enlarged compartment(s) to maintain a microbiota and other GI structures that slow digesta flow. The taxon richness of the gut microbiota, usually identified by 16S rRNA gene sequencing, is typically an order of magnitude greater in vertebrates than invertebrates, and the interspecific variation in microbial composition is strongly influenced by diet. Many of the nutrient transporters are orthologous across different animal phyla, though functional details may vary (e.g., glucose and amino acid transport with K+ rather than Na+ as a counter ion). Paracellular absorption is important in many birds. Natural toxins are ubiquitous in foods and may influence key features such as digesta transit, enzymatic breakdown, microbial fermentation, and absorption.
在脊椎动物和无脊椎动物中,胃肠道(GI)的形态和功能特征通常反映食物化学性质,例如碳水化合物、蛋白质、脂肪的含量和不易快速消化的物质(例如纤维素)。消化酶和营养转运蛋白的表达大致与各自底物的饮食负荷相匹配,具有相对适度的过剩能力。解释种群和物种之间水解酶活性差异的机制包括基因拷贝数变异和单核苷酸多态性。转录和转录后调节介导水解酶和转运蛋白表达的表型变化,以响应饮食信号。许多物种通过灵活地增加消化腔室的大小来响应更高的食物摄入量。共生微生物的发酵过程对于纤维素降解很重要,但相对较慢,因此依赖这些过程的动物通常具有特殊的扩大腔室,以维持微生物群和其他减缓食糜流动的 GI 结构。肠道微生物群的分类丰富度,通常通过 16S rRNA 基因测序来识别,在脊椎动物中通常比无脊椎动物高一个数量级,微生物组成的种间变异强烈受饮食影响。许多营养转运蛋白在不同的动物门之间是同源的,尽管功能细节可能有所不同(例如,葡萄糖和氨基酸转运与 K+而不是 Na+作为反离子)。旁细胞吸收在许多鸟类中很重要。天然毒素在食物中普遍存在,可能影响关键特征,如食糜转运、酶解、微生物发酵和吸收。