Department of Biology, McMaster University, Hamilton, ON L8S 4K1 Canada.
G3 (Bethesda). 2013 Aug 7;3(8):1363-74. doi: 10.1534/g3.113.006999.
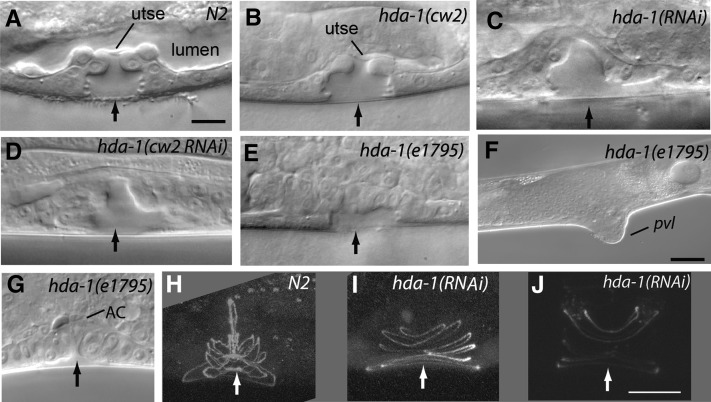
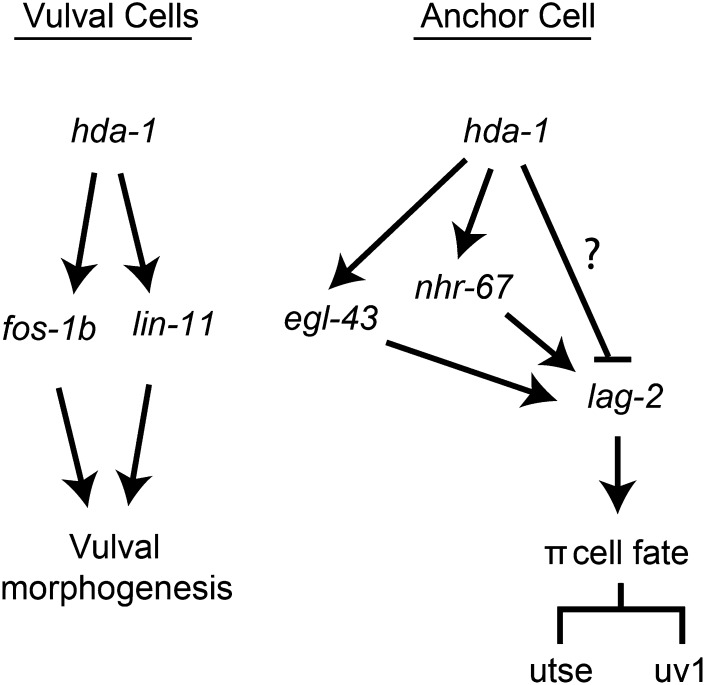
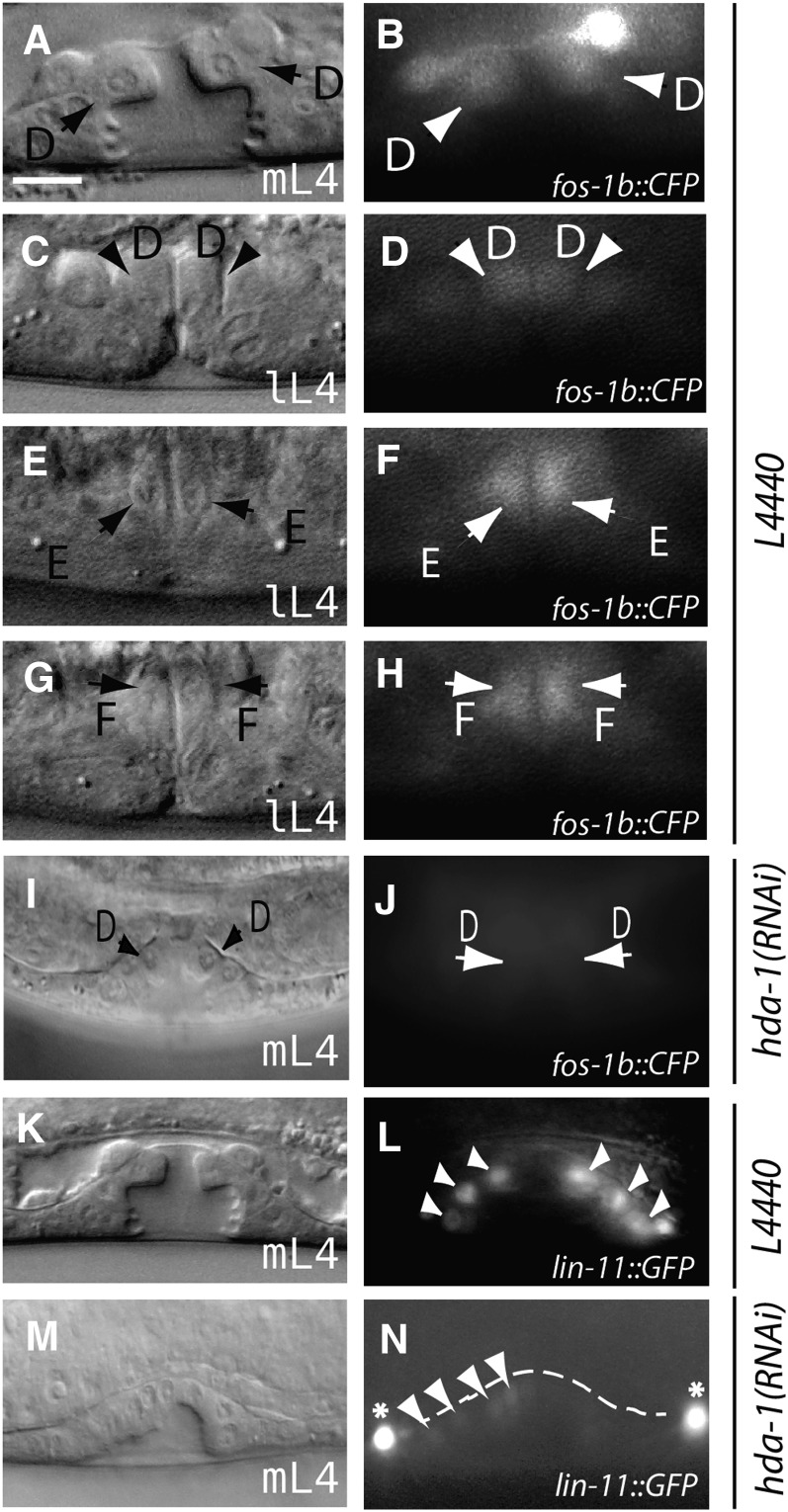
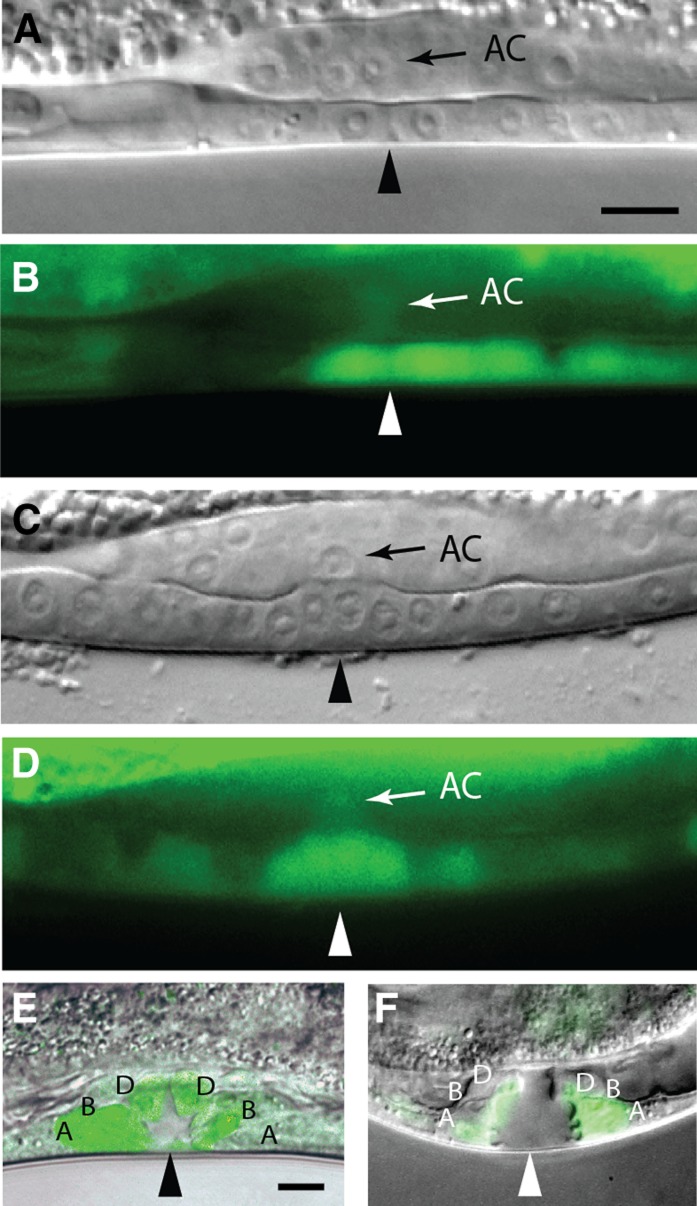
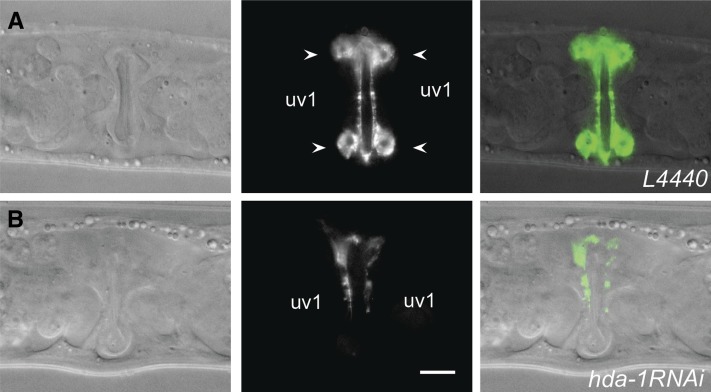
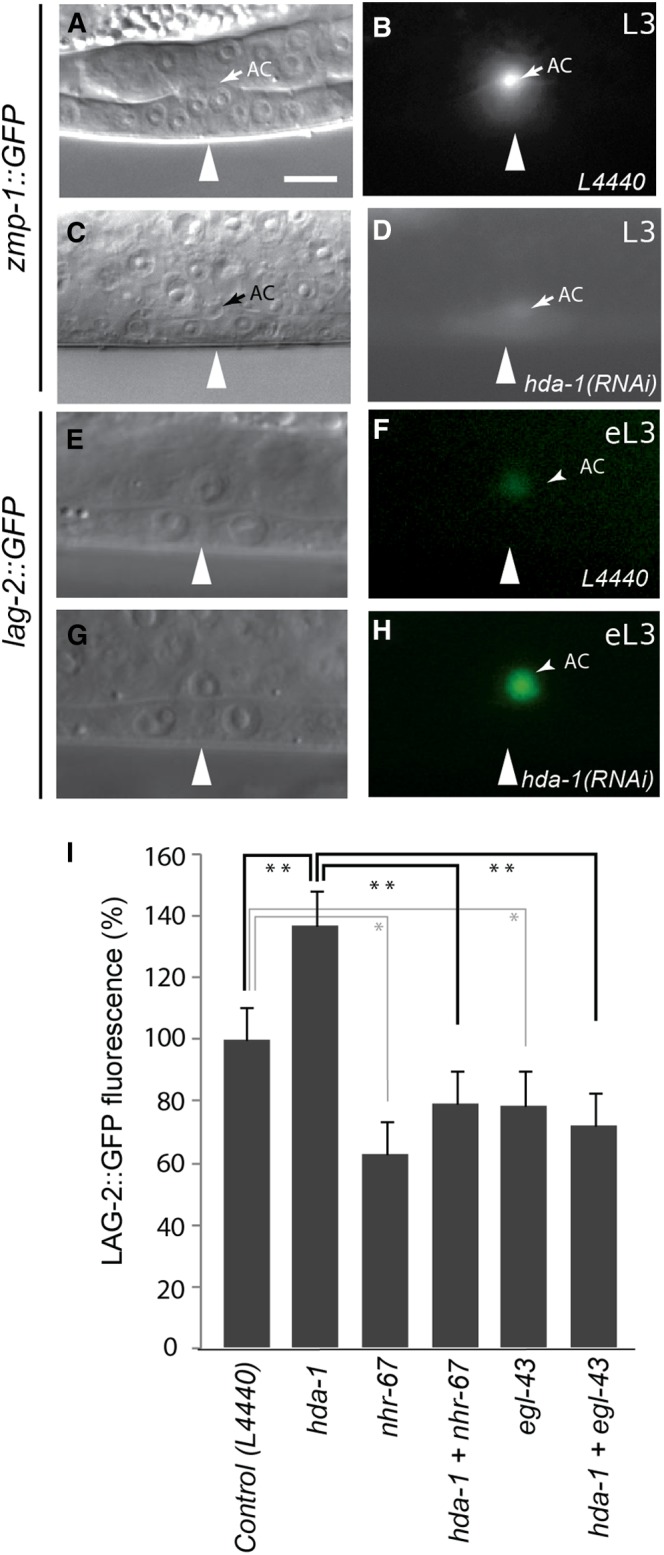
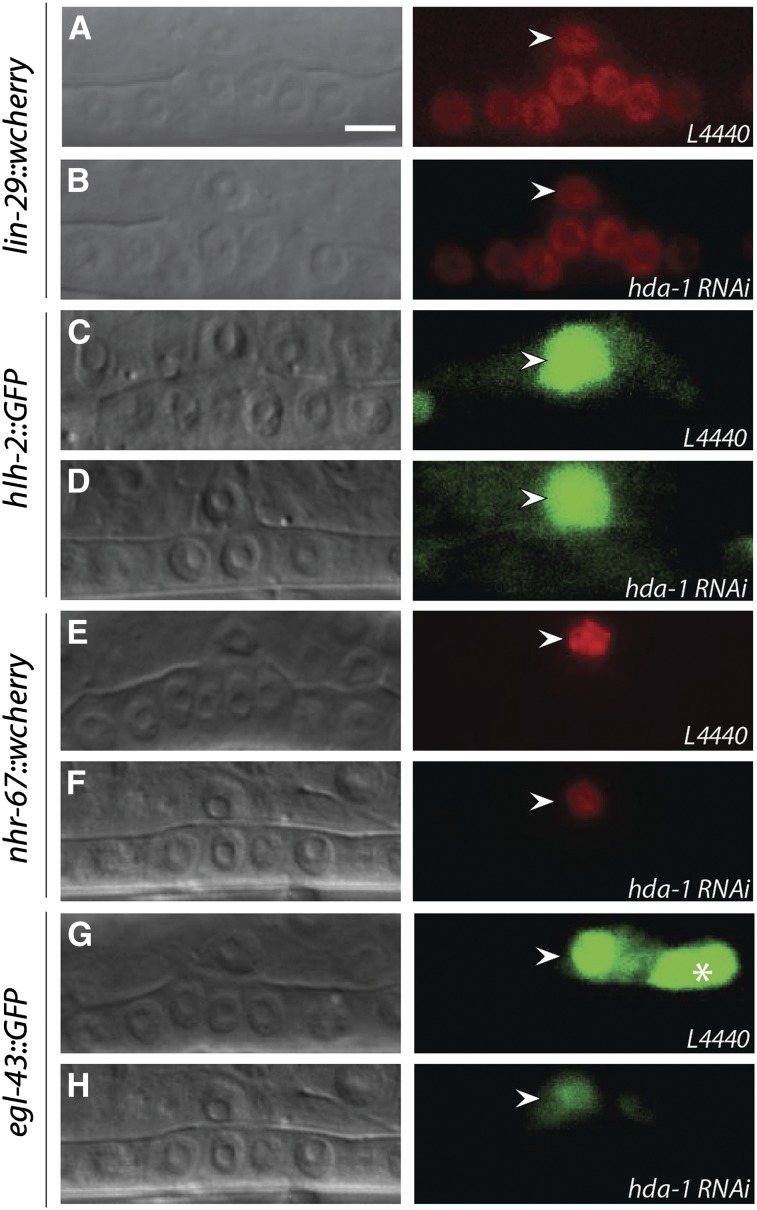
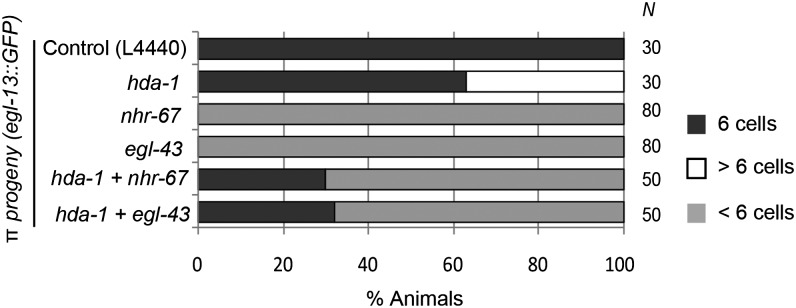
Chromatin modification genes play crucial roles in development and disease. In Caenorhabditis elegans, the class I histone deacetylase family member hda-1, a component of the nucleosome remodeling and deacetylation complex, has been shown to control cell proliferation. We recovered hda-1 in an RNA interference screen for genes involved in the morphogenesis of the egg-laying system. We found that hda-1 mutants have abnormal vulva morphology and vulval-uterine connections (i.e., no uterine-seam cell). We characterized the vulval defects by using cell fate-specific markers and found that hda-1 is necessary for the specification of all seven vulval cell types. The analysis of the vulval-uterine connection defect revealed that hda-1 is required for the differentiation of the gonadal anchor cell (AC), which in turn induces ventral uterine granddaughters to adopt π fates, leading to the formation of the uterine-seam cell. Consistent with these results, hda-1 is expressed in the vulva and AC. A search for hda-1 target genes revealed that fos-1 (fos proto-oncogene family) acts downstream of hda-1 in vulval cells, whereas egl-43 (evi1 proto-oncogene family) and nhr-67 (tailless homolog, NHR family) mediate hda-1 function in the AC. Furthermore, we showed that AC expression of hda-1 plays a crucial role in the regulation of the lin-12/Notch ligand lag-2 to specify π cell fates. These results demonstrate the pivotal role of hda-1 in the formation of the vulva and the vulval-uterine connection. Given that hda-1 homologs are conserved across the phyla, our findings are likely to provide a better understanding of HDAC1 function in development and disease.
染色质修饰基因在发育和疾病中起着至关重要的作用。在秀丽隐杆线虫中,组蛋白去乙酰化酶家族成员 hda-1 是核小体重塑和去乙酰化复合物的一个组成部分,它被证明可以控制细胞增殖。我们从涉及产卵系统形态发生的基因 RNAi 筛选中回收了 hda-1。我们发现 hda-1 突变体的外生殖器形态异常,且与子宫的连接(即没有子宫-缝线细胞)也异常。我们利用细胞命运特异性标记物对这些外生殖器缺陷进行了特征描述,发现 hda-1 对于所有七种外生殖器细胞类型的特化都是必需的。对外生殖器-子宫连接缺陷的分析表明,hda-1 对于性腺锚定细胞(AC)的分化是必需的,而性腺锚定细胞又会诱导 ventral uterine granddaughters 获得 π 命运,从而形成子宫-缝线细胞。这些结果与 hda-1 在 AC 中表达一致。对 hda-1 靶基因的搜索显示,fos-1(fos 原癌基因家族)在 vulval 细胞中是 hda-1 的下游,而 egl-43(evi1 原癌基因家族)和 nhr-67(tailless 同源物,NHR 家族)介导 hda-1 在 AC 中的功能。此外,我们还表明,AC 中 hda-1 的表达对于调节 lin-12/Notch 配体 lag-2 以特化 π 细胞命运起着关键作用。这些结果表明 hda-1 在形成外生殖器和外生殖器-子宫连接中起着关键作用。鉴于 hda-1 同源物在门之间是保守的,我们的发现可能为更好地理解 HDAC1 在发育和疾病中的功能提供依据。