Nybom Hilde, Weising Kurt, Rotter Björn
Plant Molecular Systematics, Institute of Biology, University of Kassel, Kassel 34109, Germany.
Investig Genet. 2014 Jan 3;5(1):1. doi: 10.1186/2041-2223-5-1.
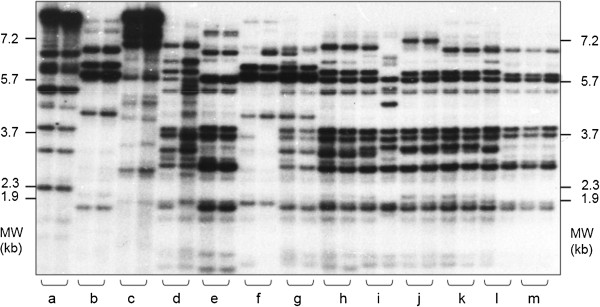
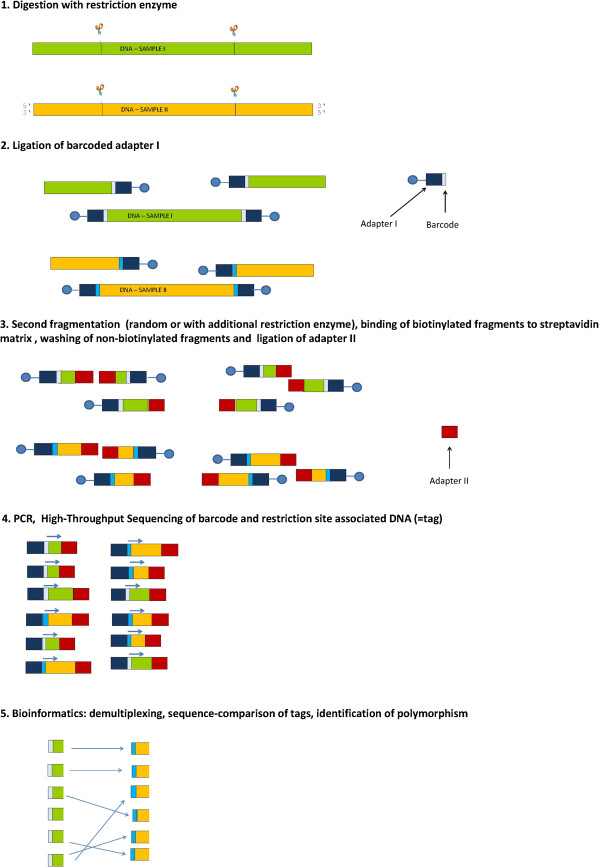
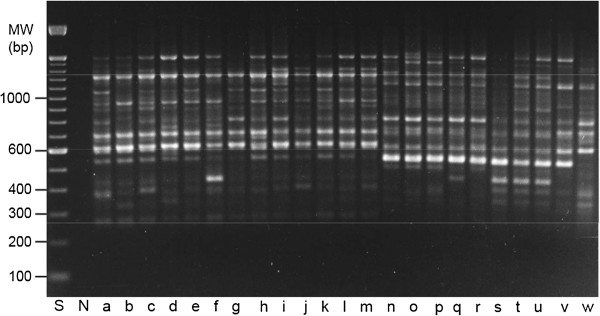
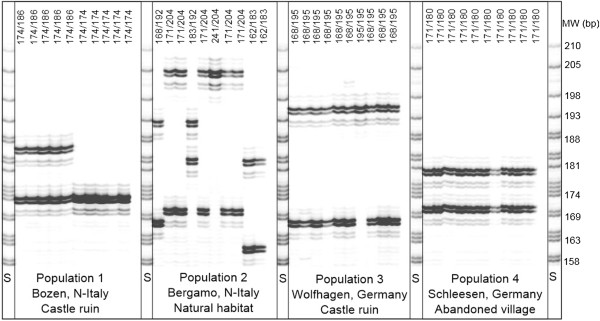
Almost three decades ago Alec Jeffreys published his seminal Nature papers on the use of minisatellite probes for DNA fingerprinting of humans (Jeffreys and colleagues Nature 1985, 314:67-73 and Nature 1985, 316:76-79). The new technology was soon adopted for many other organisms including plants, and when Hilde Nybom, Kurt Weising and Alec Jeffreys first met at the very First International Conference on DNA Fingerprinting in Berne, Switzerland, in 1990, everybody was enthusiastic about the novel method that allowed us for the first time to discriminate between humans, animals, plants and fungi on the individual level using DNA markers. A newsletter coined "Fingerprint News" was launched, T-shirts were sold, and the proceedings of the Berne conference filled a first book on "DNA fingerprinting: approaches and applications". Four more conferences were about to follow, one on each continent, and Alec Jeffreys of course was invited to all of them. Since these early days, methodologies have undergone a rapid evolution and diversification. A multitude of techniques have been developed, optimized, and eventually abandoned when novel and more efficient and/or more reliable methods appeared. Despite some overlap between the lifetimes of the different technologies, three phases can be defined that coincide with major technological advances. Whereas the first phase of DNA fingerprinting ("the past") was dominated by restriction fragment analysis in conjunction with Southern blot hybridization, the advent of the PCR in the late 1980s gave way to the development of PCR-based single- or multi-locus profiling techniques in the second phase. Given that many routine applications of plant DNA fingerprinting still rely on PCR-based markers, we here refer to these methods as "DNA fingerprinting in the present", and include numerous examples in the present review. The beginning of the third phase actually dates back to 2005, when several novel, highly parallel DNA sequencing strategies were developed that increased the throughput over current Sanger sequencing technology 1000-fold and more. High-speed DNA sequencing was soon also exploited for DNA fingerprinting in plants, either in terms of facilitated marker development, or directly in the sense of "genotyping-by-sequencing". Whereas these novel approaches are applied at an ever increasing rate also in non-model species, they are still far from routine, and we therefore treat them here as "DNA fingerprinting in the future".
大约三十年前,亚历克·杰弗里斯在《自然》杂志上发表了具有开创性的论文,介绍了利用小卫星探针进行人类DNA指纹识别的方法(杰弗里斯及其同事,《自然》1985年,第314卷:67 - 73页;《自然》1985年,第316卷:76 - 79页)。这项新技术很快被应用于包括植物在内的许多其他生物。1990年,希尔德·尼博姆、库尔特·魏辛和亚历克·杰弗里斯在瑞士伯尔尼举行的第一届国际DNA指纹识别会议上首次见面时,每个人都对这种新技术充满热情,它首次使我们能够利用DNA标记在个体水平上区分人类、动物、植物和真菌。一份名为《指纹新闻》的时事通讯创刊了,还出售了相关T恤,伯尔尼会议的论文集成为了第一本关于《DNA指纹识别:方法与应用》的书籍。随后还将举办另外四场会议,每个大洲各一场,亚历克·杰弗里斯当然受邀参加了所有会议。从早期开始,方法就经历了快速的演变和多样化。众多技术被开发、优化,当更新颖、更高效和/或更可靠的方法出现时,一些技术最终被淘汰。尽管不同技术的使用期限存在一些重叠,但可以定义三个与主要技术进步相契合的阶段。DNA指纹识别的第一阶段(“过去”)以限制性片段分析结合Southern印迹杂交为主,20世纪80年代末PCR的出现为第二阶段基于PCR的单基因座或多基因座分析技术的发展铺平了道路。鉴于植物DNA指纹识别的许多常规应用仍然依赖基于PCR的标记,我们在此将这些方法称为“当前的DNA指纹识别”,并在本综述中纳入了大量实例。第三阶段实际上始于2005年,当时开发了几种新颖的、高度并行的DNA测序策略,使通量比当前的桑格测序技术提高了1000倍甚至更多。高速DNA测序很快也被用于植物的DNA指纹识别,要么用于促进标记开发,要么直接用于“测序基因分型”。尽管这些新方法在非模式物种中的应用率也在不断提高,但它们仍远未成为常规方法,因此我们在此将它们视为“未来的DNA指纹识别”。